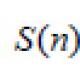Как называется симбиоз гриба водоросли. Симбиоз в растительном мире
- Гриб- поглощает минеральные вещества, выделяет углекислоту и воду (для водоросли), вырабатывает ряд веществ стимклирующих развитие водоросли.
- просвятительские
- Симбиотические. Больше у меня нет слов:)
Водоросль- вырабатывает улеводы, которые потребляет гриб.
В результате имеем "взаимовыгодное сотрудничество"- симбиоз
Существует несколько теорий, объясняющих взаимоотношения и водоросли в лишайниках, хотя еще не - biofine.ru
Практическое значение лишайников состоит в том, что они используются для медицинских препаратов, красителей, в парфюмерной промышленности как обладающие ароматическими свойствами. Они служат индикаторами загрязнения воздуха, имеют определенное кормовое значение, особенно для северных оленей. Съедобны также некоторые лишайники, произрастающие в степной и пустынной зонах, например Aspicilia esculenta, содержащий до 55-65 % оксалата кальция. У лишайника Romalina duriaci, произрастающего на нижних мертвых ветвях деревьев Acacia tortilis, белка составляет 7,4 %, а углеводы составляют более половины - 55,4 % массы лишайника, в том числе усвояемых - 28,7 %.
В литературе описана также ассоциация лишайника Usnea strigosa с насекомыми Lanelognatha theraiis, которая, видимо, строится на биологической роли лишайниковых кислот.
Взаимоотношения гриба и водоросли в теле лишайника
Отдел лишайники
Отдел лишайники занимают особое место в растительном мире. Их строение очень своеобразно. Тело, называемое слоевищем, состоит из двух организмов — гриба и водоросли, живущих как один организм, В составе некоторых видов лишайников обнаружены бактерии. Такие лишайники представляют собой тройной симбиоз.
Слоевище образовано переплетением гиф гриба с клетками водорослей (зеленых и сине-зеленых).
срез тела листоватого лишайника" width="489" height="192" title="Поперечный срез тела листоватого лишайника" />
Живут лишайники на скалах, деревьях, почве, как на Севере, так и в тропических странах. Разные виды лишайников имеют различную окраску - от серой, желтоватой, зеленоватой до бурой и черной. В настоящее время известно более 20 000 видов лишайников. Изучает лишайники наука, которая называется лихенологией (от греч. «лейхен» - лишайник и «логос» - наука).
По морфологическим признакам (внешнему виду) лишайники делятся на три группы.
- Накипные, или корковые, прикрепляющиеся к субстрату очень плотно, образуя корку. Эта группа составляет около 80% всех лишайников.
- Листоватые, представляющие собой пластинку, похожую на пластинку листа, слабо прикрепленную к субстрату.
- Кустистые, представляющие собой свободные маленькие кустики.
Лишайники - очень неприхотливые растения. Они в самых бесплодных местах. Их можно встретить на голых скалах, высоко в горах, где не живут другие растения. Растут лишайники очень медленно. Например, «олений мох» (ягель) за год вырастает всего на 1 - 3 мм. Живут лишайники до 50 лет, а некоторые до 100 лет.
Размножаются лишайники вегетативно, кусочками слоевища, а также особыми группами клеток, появляющихся внутри их тела. Эти группы клеток образуются во множестве. Тело лишайника разрывается под давлением их разросшейся массы, и группы клеток разносятся ветром и дождевыми потоками.
Лишайники в природе и в хозяйственной деятельности играют важную роль. Лишайники являются первыми растениями, которые поселяются на скалах и им подобных бесплодных местах, где другие растения жить не могут. Лишайники разрушают поверхностный слой скалы и, отмирая, образуют слой гумуса, на котором уже могут поселяться другие растения.
Значение для жизнедеятельности лишайников
Чаще всего в качестве неверного ответа указывают, что грибы, входящие в лишайника, обеспечивают половое размножение водоросли.
Обмен веществ у лишайников также особенный, не сходный ни с водорослями, ни с грибами. Лишайники образуют особые вещества, больше нигде в природе не встречающиеся. Это лишайниковые кислоты . Некоторые из них обладают стимулирующим, или антибиотическим, действием, например, усниновая кислота. Вероятно, поэтому ряд лишайников издавна применялся в народной медицине как противовоспалительное, вяжущее или тонизирующее средство - отвары «исландского мха», например.
Благодаря сочетанию в одном организме гриба и водоросли лишайники обладают рядом уникальных свойств.
Во-первых , это их способность расти там, где никакое другое растение не может поселиться и выжить: на камнях и скалах в самых суровых условиях Арктики или высокогорий, на беднейших почвах тундр, торфяных болотах, на песках, на таких малопригодных для жизни предметах, как стекло, железо, кирпичи, черепица, кости. Лишайники находили на смоле, фаянсе, фарфоре, коже, картоне, линолеуме, древесном угле, войлоке, полотняных и шелковых тканях и даже на старинных пушках! Именно лишайники первыми осваивают непригодную для других организмов среду обитания, например вулканические лавы, разлагая их. За это лишайники получили название «пионеров растительности» Они прокладывают дорогу другим растениям. Вслед за лишайниками поселяются мхи и зеленые травянистые растения Лишайники легко переносят пятидесятиградусные морозы в тундре, а в пустынях Азии и Африки - шестидесятиградусную жару. Легко переносят они и сильное высыхание.
Вторая особенность лишайников - их крайне медленный рост. Ежегодно лишайник вырастает на один-пять миллиметров. Необходимо оберегать лишайниковый покров тундры, хвойных боров. Если его нарушить, он восстанавливается очень долго. маленький срок - около десяти лет. Лишенный такого покрова, тонкий слой почвы в тундре или сосняках подвергается эрозии, а это ведет к гибели и другой растительности.
Средний возраст лишайников от тридцати до восьмидесяти лет, а отдельные экземпляры, как это удалось установить по косвенным данным, доживают до шестисот лет. Имеются сведения, что некоторые лишайники насчитывают даже около двух тысяч лет. Наряду с секвойей и остистой сосной лишайники можно считать самыми долгоживущими организмами.
Лишайники очень чувствительны к чистоте окружающего воздуха . Если в воздухе содержится значительная концентрация углекислого и особенно сернистого газа, лишайники исчезают. Эту их особенность предлагается использовать для оценки чистоты воздуха в городах и промышленных районах.
Своеобразие формы тела, обмена веществ, особенностей роста, мест обитания позволяет считать лишайники, несмотря на их двойственную природу, самостоятельными организмами.
Симбиоз гриба и водоросли
Итак, в лабораториях, в стерильных пробирках и колбах с питательной средой поселились изолированные симбионты лишайников.Имея в распоряжении чистые культуры лишайниковых партнеров, ученые решились на самый дерзкий шаг - синтез лишайника в лабораторных условиях.Первая удача на этом поприще принадлежит Е. Томасу, который в 1939 году в Швейцарии получил из мико- и фотобионтов лишайник кладония крыночковидная с хорошо различимыми плодовыми телами. В отличие от предыдущих исследователей, Томас выполнял синтез в стерильных условиях, что внушает доверие к полученному им результату. К сожалению, его попытки повторить синтез в 800 других опытах не удались.
Любимый объект исследования В. Ахмаджяна, принесший ему всемирную славу в области лишайникового синтеза, - кладония гребешковая. Этот лишайник широко распространен в Северной Америке и получил простонародное название "британские солдаты": его ярко-красные плодовые тела напоминают алые мундиры английских солдат времен войны североамериканских колоний за независимость.Небольшие комочки изолированного микобионта кладонии гребешковой смешивали с фотобионтом, извлеченным из того же лишайника. Смесь помещали на узкие слюдяные пластинки, пропитанные минеральным питательным раствором и закрепленные в закрытых колбах. Внутри колб поддерживали строго контролируемые условия влажности, температуры и освещенности. Важным условием эксперимента было минимальное количество питательных веществ в среде. Как же вели себя лишайниковые партнеры в непосредственной близости друг к другу? Клетки водоросли выделяли особое вещество, которое "приклеивало" к ним гифы гриба, и гифы сразу начинали активно оплетать зеленые клетки. Группы водорослевых клеток скреплялись ветвящимися гифами в первичные чешуйки. Следующим этапом было дальнейшее развитие утолщенных гиф поверх чешуек и выделение ими внеклеточного материала, а в результате - образование верхнего корового слоя. Еще позже дифференцировались водорослевый слой и сердцевина, совсем как в слоевище природного лишайника. Эти опыты были многократно воспроизведены в лаборатории Ахмаджяна и всякий раз приводили к появлению первичного лишайникового слоевища.
В 40-е годы XX века немецкий ученый Ф. Тоблер обнаружил, что для прорастания спор ксантории настенной требуются добавки стимулирующих веществ: экстрактов из древесной коры, водорослей, плодов сливы, некоторых витаминов или других соединений. Было сделано предположение, что в природе прорастание некоторых грибов стимулируется веществами, поступающими из водоросли.
Примечательно, что для возникновения симбиотических отношений оба партнера получать умеренное и даже скудное питание, ограниченные влажность и освещение. Оптимальные условия существования гриба и водоросли отнюдь не стимулируют их воссоединение. Более того, известны случаи, когда обильное питание (например, при искусственном удобрении) вило к быстрому росту водорослей в слоевище, нарушению связи между симбионтами и гибели лишайника.
Если рассматривать срезы лишайникового слоевища под микроскопом, видно, что чаще всего водоросль просто соседствует с грибными гифами. Иногда гифы тесно прижимаются к водорослевым клеткам. Наконец, грибные гифы либо их ответвления могут более или менее глубоко проникать внутрь водоросли. Эти выросты называются гаусториями.
Совместное существование накладывает отпечаток и на строение обоих лишайниковых симбионтов. Так, если свободноживущие синезеленые водоросли родов носток, сцитонема и других образуют длинные, иногда ветвящиеся нити, то у тех же водорослей в симбиозе нити либо скручены в плотные клубочки, либо укорочены до единичных клеток. Кроме того, у свободноживущих и лихенизированных синезеленых водорослей отмечают различия в размерах и расположении клеточных структур.Зеленые водоросли также изменяются в симбиотическом состоянии. Это, в первую очередь, касается их размножения. Многие из зеленых водорослей, живя "на свободе", размножаются подвижными тонкостенными клеточками - зооспорами. В слоевище зооспоры, обычно, не образуются. Вместо них появляются апланоспоры - относительно маленькие клетки с толстыми стенками, хорошо приспособленные к засушливым условиям. Из клеточных структур зеленых фотобионтов наибольшим изменениям подвергается оболочка. Она тоньше, чем у тех же водорослей "на воле", и имеет ряд биохимических различий. Очень часто внутри симбиотических клеток наблюдают жироподобные зернышки, которые после изъятия водоросли из слоевища исчезают. Говоря о причинах этих различий, можно предположить, что они связаны с каким-то химическим воздействием грибного соседа водоросли.Сам микобионт также испытывает воздействие водорослевого партнера. Плотные комочки изолированных микобионтов, состоящие из тесно переплетенных гиф, внешне совсем не похожи на лихенизированные грибы. Внутреннее строение гиф тоже различно. Клеточные стенки гиф в симбиотическом состоянии значительно тоньше.
Итак, жизнь в симбиозе побуждает водоросль и гриб менять свой внешний облик и внутреннее строение.
Что же получают сожители друг от друга, какую пользу извлекают из совместного существования? Водоросль снабжает гриб, своего соседа по лишайниковому симбиозу, углеводами, полученными в процессе фотосинтеза.Водоросль, синтезировав тот или иной углевод, быстро и почти целиком отдает его своему грибному "сожителю". Гриб получает от водоросли не только углеводы. Если синезеленый фотобионт фиксирует атмосферный азот, существует быстрый и устойчивый отток образовавшегося аммония к грибному соседу водоросли. Водоросль же, очевидно, просто получает возможность широко расселяться по Земле. По словам Д. Смита, "наиболее частая у лишайников водоросль, требуксия, очень редко живет вне лишайника. Внутри же лишайника она распространена, пожалуй, шире, чем любой род свободноживущих водорослей. за занятие этой ниши - снабжение гриба-хозяина углеводами".
Литература
Лишайники - википедия
Биохимические особенности[править]
Большинство внутриклеточных продуктов, как фото-(фико-), так и микобионтов не являются специфичными для лишайников. Уникальные вещества (внеклеточные), так называемые лишайниковые , формируются исключительно микобионтом и накапливаются в его гифах. Сегодня известно более 600 таких веществ, например, усниновая кислота, мевалоновая кислота. Нередко, именно эти вещества оказываются решающими в формировании окраски лишайника. Лишайниковые кислоты играют важную роль в выветривании, разрушая субстрат.
Водный обмен[править]
Лишайники не способны к регуляции водного баланса, поскольку у них нет настоящих корней для активного поглощения воды и защиты от испарения. Поверхность лишайника может удерживать воду на короткое время в форме жидкости или пара. В условиях вода быстро теряется на поддержание метаболизма и лишайник переходит в фотосинтетически неактивное состояние, при котором вода может составлять не более 10 % массы. В отличие от микобионта, фотобионт не может долго находиться без воды. Сахар трегалоза играет важную роль в защите жизненно важных макромолекул, таких как ферменты, мембранные элементы и ДНК. Но лишайники нашли способы предотвращения полной потери влаги. У многих видов наблюдается утолщение коры, чтобы обеспечить меньшую потерю воды. Способность поддерживать воду в жидком состоянии очень важна в холодных районах, поскольку замёрзшая вода не пригодна для использования организмом.
Время, которое лишайник может провести высушенным, зависит от вида, известны случаи «воскрешения» после 40 лет в сухом состоянии. Когда поступает пресная вода в форме дождя, росы или влажности, лишайники быстро переходят в активное состояние, возобновляя метаболизм. Оптимально для жизнедеятельности, когда вода составляет от 65 до 90 процентов от массы лишайника. Влажность в течение дня может изменяться в зависимости от темпов фотосинтеза, как правило, она наиболее высока с утра, когда лишайники смачиваются росой.
Рост и продолжительность жизни[править]
Описанный выше ритм жизни является одной из причин для очень медленного роста большинства лишайников. Иногда лишайники растут всего лишь на несколько десятых миллиметра в год, в основном менее чем на один сантиметр. Другой причиной медленного роста является то, что фотобионт, составляя нередко менее 10 % объёма лишайника, берёт на себя обеспечение микобионта питательными веществами. В хороших условиях, с оптимальными влажностью и температурой, например в туманных или дождливых тропических лесах, лишайники растут на несколько сантиметров в год.
Ростовая зона лишайников у накипных форм находится по краю лишайника, у листоватых и кустистых - на каждой верхушке.
Лишайники являются одними из самых долгоживущих организмов и могут достигать возраста нескольких сотен лет, а в некоторых случаях - более 4500 лет, как например Rhizocarpon geographicum , живущий в Гренландии.
Размножение[править]
Лишайники размножаются вегетативным, бесполым и половым путём.
Особи микобионта размножаются всеми способами и в то время, когда фотобионт не размножается или размножается вегетативно. Микобионт может, как и другие грибы, также размножаться половым и собственно бесполым путем. Половые споры в зависимости от того, относится микобионт к сумчатым или базидиальным грибам, называются аско- или базидиоспорами и образуются соответственно в асках (сумках) или базидиях .
Считается, что мутуализм (взаимовыгодный симбиоз) двух видов живых существ должен формироваться постепенно, в результате долгой коэволюции. Однако эксперименты американских биологов показали, что многие виды грибов и одноклеточных водорослей могут образовывать мутуалистические системы практически мгновенно, без предшествующего периода взаимной адаптации и без каких-либо генетических модификаций. Для этого гриб и водоросль должны оказаться в среде, где они будут друг для друга единственными источниками необходимых веществ, таких как углекислый газ и аммоний. Исследование подтвердило гипотезу «экологического соответствия», согласно которой не все существующие в природе мутуалистические системы следует трактовать как результат длительной предшествующей коэволюции.
Облигатным (обязательным) мутуализмом называют взаимовыгодные отношения между двумя видами, не способными существовать друг без друга. Принято считать, что такие отношения формируются постепенно, в ходе длительной коэволюции и взаимной адаптации, «притирки» организмов друг к другу. Несомненно, во многих случаях так оно и было (см. Н. Проворов, Е. Долгих, 2006. Метаболическая интеграция организмов в системах симбиоза).
Разумеется, не всякий вид способен встроиться в новое окружение. При интродукции происходит своеобразная сортировка, в ходе которой одни пришельцы приживаются на новом месте, а другие погибают. Так или иначе, приходится признать, что целостное и взаимосвязанное сообщество может сформироваться не только за счет идущей миллионы лет коэволюционной «притирки» видов друг к другу, но и за счет подбора из числа случайных мигрантов таких видов, которые удачно дополняют друг друга и хорошо уживаются вместе. Эту идею, известную под названием ecological fitting (что можно приблизительно перевести как «экологическое соответствие» или «экологический подбор»), начиная с 1980-х годов развивает известный американский эколог Дэниел Джензен (Daniel Janzen).
Могут ли облигатно-мутуалистические системы, обычно считающиеся чем-то вроде апофеоза коэволюции, формироваться по такой же схеме, то есть без всякой коэволюции - просто за счет случайного соответствия двух случайно встретившихся видов, которые при определенных условиях оказываются неспособными жить друг без друга? Эксперименты, проведенные биологами из Гарвардского университета (США), позволяют ответить на этот вопрос утвердительно.
Авторы работали с обычными пекарскими почкующимися дрожжами Saccharomyces cerevisiae и не менее обычными одноклеточными водорослями хламидомонадами (Chlamydomonas reinhardtii). В природе эти виды в мутуалистических отношениях замечены не были. В лаборатории, однако, они вступили в неразрывную связь легко и быстро, без всякой эволюции или генетических модификаций. Для этого оказалось достаточно выращивать дрожжи и хламидомонады без доступа воздуха в среде, где глюкоза является единственным источником углерода, а нитрит калия - единственным источником азота.
Схема мутуалистических взаимоотношений дрожжей и хламидомонад довольно проста (рис. 1). Дрожжи питаются глюкозой и производят углекислый газ, необходимый хламидомонадам для фотосинтеза (использовать содержащуюся в среде глюкозу хламидомонады не умеют). Водоросли, со своей стороны, восстанавливают нитрит, переводя азот в доступную для дрожжей форму (аммоний). Таким образом, дрожжи обеспечивают хламидомонады углеродом, а хламидомонады снабжают дрожжи азотом. В таких условиях ни один из видов не может расти без другого. Это и есть облигатный мутуализм.
Авторы убедились, что мутуалистическая система благополучно растет в широком диапазоне концентраций глюкозы и нитрита, хотя в одиночку ни один из двух видов в этих условиях не выживает. Только при очень сильном снижении концентрации глюкозы или нитрита рост смешанной культуры прекращается.
Если раскупорить систему, то есть предоставить ей доступ к атмосферному CO2, получается сообщество, в котором только один из участников (дрожжи) не может жить без другого, тогда как второй участник (хламидомонады) уже не нуждается в первом для выживания. Впрочем, даже в этом случае хламидомонады лучше растут в присутствии дрожжей, чем без них (очевидно, дополнительный CO2, выделяемый дрожжами, идет им на пользу). Таким образом, система остается мутуалистической, хотя со стороны водорослей мутуализм уже не облигатный. Ни один из видов не вытесняет другой.
Если добавить в среду аммоний, получается обратная ситуация: теперь дрожжи могут жить без водорослей (и вообще не нуждаются в них), тогда как водоросли по-прежнему не могут жить без дрожжей. Это уже не мутуализм, а комменсализм (нахлебничество со стороны водорослей). В этом случае дрожжи, которые размножаются быстрее водорослей, заполняют всё жизненное пространство, доводя хламидомонады до вымирания. Авторы предполагают, что устойчивость таких асимметричных систем (в которых только один из участников сильно зависит от другого) определяется соотношением скоростей размножения. Если зависимый вид размножается быстрее, чем независимый, то сожительство двух видов может быть устойчивым; в противном случае независимый вид может полностью вытеснить своего напарника.
Авторы провели аналогичные эксперименты с другими видами хламидомонад и грибов-аскомицетов. Оказалось, что почти все виды дрожжей в данных условиях образуют облигатно-мутуалистические взаимоотношения с хламидомонадами. Правда, продуктивность (скорость роста) симбиотических комплексов оказывается разной. От чего она зависит, определить не удалось: авторы не нашли связи ни со склонностью дрожжей к кислородному дыханию или бескислородному метаболизму (брожению), ни с природными местообитаниями дрожжей, ни со скоростью размножения, ни со степенью влияния концентрации нитритов на рост дрожжей. Очевидно, дело в каких-то других особенностях изученных видов.
Одноклеточная водоросль хлорелла отказалась вступать в мутуалистические отношения с дрожжами, потому что она сама умеет питаться глюкозой и в смешанной культуре вытесняет дрожжи. Не стали образовывать облигатно-мутуалистические комплексы с водорослями дрожжи Hansenula polymorpha, потому что они сами умеют использовать нитрит в качестве источника азота. Но все же исследование показало, что самые разные виды аскомицетов и хламидомонад готовы вступить в симбиотические отношения друг с другом, попав в подходящие условия.
Из многоклеточных (точнее, образующих нитчатые гифы) аскомицетов были протестированы два классических лабораторных объекта - Neurospora crassa и Aspergillus nidulans. Оба вида умеют восстанавливать нитрит и потому не образуют облигатно-мутуалистических систем с хламидомонадами. Однако генетически модифицированные штаммы этих грибов, лишенные способности утилизировать нитрит, вступили в симбиоз с водорослями точно так же, как и дрожжи. Как выяснилось, при этом клетки хламидомонад вступают в непосредственный физический контакт с гифами грибов: под микроскопом видны гифы, обвешанные хламидомонадами, как новогодняя елка (рис. 2).
Мутуалистические взаимоотношения хламидомонад с дрожжами, по-видимому, тоже требуют установления физических контактов между клетками. Об этом свидетельствует тот факт, что систематическое встряхивание смешанной культуры дрожжей и водорослей резко замедляет рост симбиотической системы.
При помощи электронного микроскопа авторы обнаружили плотные контакты, образующиеся между клеточными стенками Aspergillus nidulans и Chlamydomonas reinhardtii, причем клеточная стенка водоросли в местах контакта становится тоньше - возможно, под действием ферментов, выделяемых грибом.
Похожие межклеточные контакты характерны для классических грибно-водорослевых симбиотических систем - лишайников. Аскомицеты в ходе своей эволюции много раз вступали в симбиоз с водорослями и цианобактериями, образуя лишайники. Лишайникообразующие группы разбросаны по всему филогенетическому дереву аскомицетов. Это значит, что такие эволюционные события происходили многократно и независимо в разных эволюционных линиях грибов (см. F. Lutzoni et al., 2001. Major fungal lineages are derived from lichen symbiotic ancestors). По-видимому, аскомицеты в целом «предрасположены» (преадаптированы) к формированию мутуалистических комплексов с одноклеточными водорослями. Эксперименты американских ученых, возможно, проливают свет на ранние стадии формирования таких комплексов.
Впрочем, не следует переоценивать сходство полученных в эксперименте мутуалистических систем с лишайниками. Хотя бы потому, что у большинства лишайников только грибной компонент не может жить в одиночку, тогда как фотосинтезирующие компоненты (одноклеточные водоросли и цианобактерии), как правило, могут прекрасно жить и без гриба. То есть лишайники не являются облигатно-мутуалистическими системами. Да и отсутствие доступа к атмосферному CO2 вряд ли является проблемой, с которой водорослям часто приходится сталкиваться в природе. Главное в обсуждаемой работе - демонстрация общего принципа. Исследование показало, что облигатный мутуализм может сложиться мгновенно, без всякой эволюции - просто за счет того, что изменившиеся условия делают виды взаимозависимыми. Разумеется, для того, чтобы из такого наспех сформированного симбиотического комплекса развилось что-то действительно сложное и высоко интегрированное, вроде лишайника, без миллионов лет коэволюции уже не обойтись.


Кира Столетова
Самый загадочный симбиоз гриба и водоросли – класс лишайников. Организм, состоящий из двух компонентов, исследует наука, которая называется лихенология. До сих пор ученым не удалось установить природу их возникновения, а в лабораторных условиях их получают с большим трудом.

Состав организма
Ранее думали, что в симбиоз грибов и водорослей в лишайнике представлен взаимовыгодным способом сосуществования двух организмов:
- грибы получают углеводы, производимые вторым компонентом;
- водорослям необходимы минеральные вещества и покров, чтобы защититься от засухи.
Сейчас союз представляют по-другому: споры гриба выбирают себе кормилицу, но последняя может противостоять объединению. Главное правило в симбиозе – взаимовыгодное состояние. Лишайник получится, если оба компонента испытывают трудности существования поодиночке: им не хватает питания, света, температуры. Благоприятные факторы не вынуждают их объединяться.
Вступающие во взаимодействие грибы по-разному ведут себя с водорослями. Образует гиф со всеми доступными видами, но некоторых из них просто съедают. Синтез проявляется только со схожими классами. В сосуществовании оба организма меняют свое строение и внешний вид.
Строение организма
Структурно в лишайнике представлено 2 компонента: гифы грибов с вплетенными в них водорослями. Если вплетение равномерное, его называют гомеомерным, а если только в верхнем шаре – гетеромерным. Это так называемое слоевище таллома.
Тело организма называется таллом. По внешнему виду выделяют типы:
- накипные;
- листоватые;
- кустистые.
Первые выглядят как тонкая корка, крепко сросшаяся с поверхностью. Листоватые держатся на пучках гифов. Кустистые выглядят как свисающий куст или борода.
Цвет может быть серый, бурый, зеленоватый, желтый или черный. Концентрация регулируется специфическими красящими веществами, содержанием железа, кислотами в окружающей среде.
Способы размножения и жизненный цикл

В лишайнике способностью размножаться наделены оба компонента. Грибной воспроизводится вегетативно – частями таллома или с помощью спор. Отростки тела отрываются от слани и перемещаются животными, людьми или ветром. Так же распространяются споры.
Второй компонент делится вегетативно. Симбиотический комплекс улучшает способность к размножению. А некоторые виды практически не существуют вне пределов лишайника.
Растут организмы медленно. Образует прирост на год от 0.25 до 36 мм. Зато они нетребовательны к условиям среды:
- растут на скалах, земле, стволах и ветках деревьев, на неорганике: стекле, металле;
- выдерживают обезвоживание.
Устойчивые к температурам от -47 до 80℃, 200 видов обитает в Антарктике. Около двух недель смогли прожить вне земной атмосферы.
Роль лишайников
Существует около 20 тыс. видов. Симбионт образует сетку распространения по всему миру. Особо значимы организмы в местностях тундры и лесов.
За прошедшие со времен Борне 100 лет в слоевище лишайников было открыто и описано много различных форм абсорбционных, или всасывающих, гиф гриба. Эти гифы плотно прижимаются к клетке водоросли или проникают в нее и служат, как предполагают, для передачи веществ, которые образуют водоросли в результате своей жизнедеятельности, грибному компоненту.
Гриб должен использовать лишь часть водорослей, оставляя резерв - здоровые и нормальные водоросли, содержимым которых он мог бы питаться.
Учеными были замечены любопытные защитные реакции со стороны лишайниковых водорослей. Например, одновременно с проникновением гаустория в клетку водоросли эта клетка делилась. При этом плоскость деления, как правило, проходила как раз через участок, занятый гаусторием, а образовавшиеся в результате этого процесса дочерние клетки были свободны от гаусториев. Было замечено также, что обычно гриб поражает водоросли, уже достигшие определенной стадии зрелости. В молодых растущих водорослях происходит энергичное отложение веществ в оболочке клетки и быстрое ее утолщение. Эта толстая оболочка клетки фикобионта препятствует проникновению абсорбционных органов гриба. Однако большей частью защитная реакция водорослей против активности грибного компонента очень слаба.
Однако все высказанные по этому поводу точки зрения до сих пор остаются лишь догадками и большей частью не подтверждены экспериментально: лишайники оказались очень трудным объектом для физиологических исследований. Ученые пока не научились выращивать и поддерживать в живом состоянии слоевище лишайников в искусственных условиях. Тот контакт между грибом и водорослью, который с такой легкостью достигается в природе (достаточно вспомнить многообразие лишайников!), никак не удается воспроизвести в лабораторных условиях. Наоборот, при переносе лишайников в лабораторию этот контакт легко нарушается и растение просто погибает. Время от времени появляются сообщения об удачных опытах выращивания лишайника в условиях лаборатории, но пока эти сообщения единичны и не всегда достоверны.
Одной из причин неудач подобных попыток можно считать чрезвычайно медленный рост лишайников. Лишайники - многолетние растения. Обычно возраст взрослых слоевищ, которые можно увидеть где-нибудь в лесу на стволе деревьев или на почве, составляет не менее 20-50 лет. В северных тундрах возраст некоторых кустистых лишайников рода кладония достигает 300 лет. Слоевище лишайников, имеющих вид корочки, в год дает прирост всего 0,2-0,3 мм.
Кустистые и листоватые лишайники растут несколько быстрее - в год их слоевище увеличивается на 2-3 мм. Поэтому, чтобы вырастить взрослый лишайник в лаборатории, требуется не менее 20 лет, а может быть, и вся жизнь исследователя. Трудно проводить столь долговременный эксперимент!
Вот почему физиологические особенности лишайников, в том числе взаимоотношения компонентов, как правило, изучают на культурах изолированных мико- и фикобионтов. Этот метод очень перспективен, так как позволяет ставить длительные и воспроизводимые опыты. Но, к сожалению, данные, полученные этим методом, не могут полностью отразить те процессы, которые происходят в целом слоевище лишайника.
И тем более мы не вправе считать, что в природе, в естественных условиях, в слоевищах лишайника эти процессы протекают точно так же, как в культурах изолированных симбионтов. Вот почему все теории, пытающиеся объяснить взаимоотношения компонентов лишайников, остаются пока лишь догадками.
Более успешным оказалось изучение форм контакта между гифами гриба и клетками водорослей в слоевищах лишайников. Как показали исследования с применением электронной микроскопии, в слоевище лишайников можно встретить по крайней мере пять типов контакта между грибными гифами и водорослевыми клетками (рис. 289).
Чаще всего отдельная клетка водоросли и клетка грибной гифы находятся в непосредственном контакте друг с другом. В таком случае гриб образует специальные абсорбционные, всасывающие органы, которые проникают внутрь водорослевой клетки или плотно прижимаются к ее оболочке.
В настоящее время среди абсорбционных органов гриба в слоевище лишайников различают несколько типов: гаустории, импрессории и аппрессории.
Гаустории - это боковые выросты гиф гриба, которые прорывают оболочку клетки водоросли и проникают в ее протопласт (рис. 289, 2). Обычно в клетке водоросли развивается один гаусторий, но иногда их может быть и два. В слоевище лишайника гаустории встречаются в большом количестве и существуют продолжительное время. Было замечено, что в оболочках молодых гаусториев нет отложений целлюлозы, которая могла бы затруднять обмен между клеткой водоросли и гифой гриба. Старые гаустории почти всегда одеты довольно толстым слоем целлюлозы. Различают интрацеллюлярные (внутриклеточные) и интрамембранные (внутриоболочковые) гаустории.
У более высокоорганизованных лишайников образуются только интра мембранные гаустории. Они прорывают оболочку клетки водоросли и достигают ее протопласта, но не углубляются в него, а остаются в оболочке водорослевой клетки (рис. 289, 5). Наибольшее количество интрамембранных гаусториев образуется в слоевище лишайников весной, в начале вегетационного периода. С наступлением осени они далеко отступают от протопласта водоросли.
Другой тип всасывающих органов гриба - импрессорий - тоже боковые выросты грибных гиф, но, в отличие от гаусториев, они пе прорывают оболочку клетки водоросли, а вдавливают ее внутрь (рис. 289, 6, 7). Импрессорий отмечены у очень многих лишайников, например у широко распространенной пельтигеры (Peltigera).
Интересно, что в слоевищах, произрастающих во влажных местообитаниях, импрессории почти не развиваются, у тех же видов в сухих местообитаниях они образуются в большом количестве. При длительной засухе число импрессориев также увеличивается. Предполагают, что в засушливые периоды и в сухих местообитаниях гриб, чтобы удовлетворить потребности в питании, увеличивает свою всасывающую поверхность за счет увеличения количества и размеров импрессориев.
В отличие от гаусториев и импрессориев, образованных боковыми отростками гифы, аппрессории образуются вершиной грибной гифы. Такая вершина гифы плотно прижимается снаружи к оболочке клетки водоросли, никогда не проникая ни в ее протопласт, ни в ее внутренний слой (рис. 289, 8).
По в более старых участках слоевища можно найти немало отмерших обесцвеченных клеток - гриб рано или поздно все-таки убивает водоросли.
Такой же тип контакта между гифами гриба и клетками водорослей был найден у некоторых слизистых и базидиальных лишайников.
У ряда лишайников, в слоевище которых встречаются нитчатые улотриксовые водоросли, можно наблюдать еще один тип контакта. Как правило, в таком случае нити водорослей бывают целиком покрыты грибными гифами. Причем лишь иногда гифы образуют на поверхности водорослевой пити рыхлую сетку. Чаще же они располагаются очень густо и, срастаясь своими стенками, образуют сплошной чехол. Отдельная лопасть такого лишайника имеет вид тончайшего волоса. Под микроскопом она напоминает полую трубку, стенки которой образованы сросшимися грибными гифами; внутри трубки тянется нить водоросли.

У слизистых лишайников семейства коллемовых (Collemataceae) обычно не наблюдается никакого контакта между грибными гифами и клетками водорослей. Слоевище коллемовых не имеет дифференцированной структуры: нити водоросли посток разбросаны в беспорядке среди грибных гиф по всей толще слоевища (рис. 297, 2). Никаких абсорбционных отростков в клетках водорослей обычно пе заметно; гифы гриба и нити сине-зеленой водоросли расположены друг около друга, не вступая в видимый контакт. Предполагают, что в данном случае гриб поглощает органические вещества, ассимилируемые водорослями, прямо из слизи, которая обычно окружает нити ностока. Однако более тщательное изучение этих лишайников показало, что у многих видов коллемы (Collema) в слоевище время от времени образуются специальные абсорбционные гифы, которые тесно прижимаются к одной из клеток водорослевой пити, а через некоторое время можно наблюдать отмирание этой клетки.
Описанные выше формы контакта между гифами микобионта и клетками водорослей, по всей видимости, не исчерпывают всего многообразия способов, с помощью которых гриб и водоросль в слоевище лишайников устанавливают между собой тесную связь. Исследования в этом направлении только начинаются. Можно думать, что дальнейшее изучение тончайших структур лишайникового слоевища с помощью электронного микроскопа не только даст много нового в описании физических контактов между грибным и водорослевым компонентами лишайников, но и откроет новые горизонты в понимании их взаимоотношений.
Жизнь растений: в 6-ти томах. - М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров . 1974 .
Первая удача на этом поприще принадлежит Е. Томасу , который в 1939 году в Швейцарии получил из мико- и фотобионтов лишайник кладония крыночковидная с хорошо различимыми плодовыми телами. В отличие от предыдущих исследователей, Томас проводил синтез в стерильных условиях, что внушает доверие к полученному им результату. К сожалению, его попытки повторить синтез в 800 других опытах не удались.
Любимый объект исследования В. Ахмаджяна, принесший ему всемирную славу в области лишайникового синтеза,- кладония гребешковая. Этот лишайник широко распространен в Северной Америке и получил простонародное название "британские солдаты": его ярко-красные плодовые тела и впрямь напоминают алые мундиры английских солдат времен войны североамериканских колоний за независимость.
Небольшие комочки изолированного микобионта кладонии гребешковой смешивали с фотобионтом, выделенным из того же лишайника. Смесь помещали на узкие слюдяные пластинки, пропитанные минеральным питательным раствором и закрепленные в закрытых колбах. Внутри колб поддерживали строго контролируемые условия влажности, температуры и освещенности. Важным условием эксперимента было минимальное количество питательных веществ в среде.
Как же вели себя лишайниковые партнеры в непосредственной близости друг к другу? Клетки водоросли вырабатывали особое вещество, которое "приклеивало" к ним гифы гриба, и гифы сразу начинали активно оплетать зеленые клетки. Группы водорослевых клеток скреплялись ветвящимися гифами в первичные чешуйки. Следующим этапом было дальнейшее развитие утолщенных гиф поверх чешуек и выделение ими внеклеточного материала, а в результате - образование верхнего коревого слоя. Еще позже дифференцировались водорослевый слой и сердцевина, совсем как в слоевище природного лишайника. Эти опыты были многократно повторены в лаборатории Ахмаджяна и всякий раз приводили к появлению первичного лишайникового слоевища.

Казалось бы, разгадана одна из главных загадок лишайника: как он образуется из своих составных частей. Но из дальнейших опытов выяснилось, что все не так-то просто.
Гриб, выделенный из кладонии гребешковой, помещали рядом с водорослями других лишайников. Среди них были зеленые и синезеленые фотобионты, изолированные из 13 видов лишайников, а также свободноживущие водоросли, не встречающиеся в лишайниковом симбиозе. Оказалось, что грибные гифы делают "первые шаги знакомства" одинаково, т. е. оплетают все водоросли и даже... простые стеклянные шарики диаметром 10-15 мкм ! Но следующие этапы "лихенизации" водорослей происходили уже по-разному, в зависимости от водорослевого партнера.
Позже в той же лаборатории провели синтез другого лишайника, уснеи щетинистой, и отмечали такие же тенденции. Гифы микобионта с одинаковым успехом начинали оплетать не только клетки своей (симбиртической) водоросли, но и требуксии замечательной, характерной, как мы уже знаем, для других видов лишайников. Но если своя, родная водоросль выглядела между грибными нитями здоровой и зеленой и само слоевище уже через пять месяцев напоминало уснею, то чужеродныеводоросли в окружении микобионта были бледными, желто-зелеными, да и слоевище не имело характерного для этого лишайника нитчатого строения.
Опыты по синтезу лишайников проводятся и. в Тель-авивском университете под руководством профессора М. Галун. Эта исследовательница синтезировала ксанторию настенную, поместив рядом на агаре мико- и фотобионты этого растения. Восемь-двенадцать месяцев симбионты жили бок о бок. За это время развились бледные желто-оранжевые лопасти лишайника диаметром 2-5 мм. Сформировались и апотеции со спорами. Правда, по своему анатомическому строению лопасти искусственного лишайника не были идентичны лопастям природного.