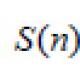Самостоятельный организм содружество гриба и водоросли. Симбиоз в растительном мире
Считается, что мутуализм (взаимовыгодный симбиоз) двух видов живых существ должен формироваться постепенно, в результате долгой коэволюции. Однако эксперименты американских биологов показали, что многие виды грибов и одноклеточных водорослей могут образовывать мутуалистические системы практически мгновенно, без предшествующего периода взаимной адаптации и без каких-либо генетических модификаций. Для этого гриб и водоросль должны оказаться в среде, где они будут друг для друга единственными источниками необходимых веществ, таких как углекислый газ и аммоний. Исследование подтвердило гипотезу «экологического соответствия», согласно которой не все существующие в природе мутуалистические системы следует трактовать как результат длительной предшествующей коэволюции.
Облигатным (обязательным) мутуализмом называют взаимовыгодные отношения между двумя видами, не способными существовать друг без друга. Принято считать, что такие отношения формируются постепенно, в ходе длительной коэволюции и взаимной адаптации, «притирки» организмов друг к другу. Несомненно, во многих случаях так оно и было (см. Н. Проворов, Е. Долгих, 2006. Метаболическая интеграция организмов в системах симбиоза).
Разумеется, не всякий вид способен встроиться в новое окружение. При интродукции происходит своеобразная сортировка, в ходе которой одни пришельцы приживаются на новом месте, а другие погибают. Так или иначе, приходится признать, что целостное и взаимосвязанное сообщество может сформироваться не только за счет идущей миллионы лет коэволюционной «притирки» видов друг к другу, но и за счет подбора из числа случайных мигрантов таких видов, которые удачно дополняют друг друга и хорошо уживаются вместе. Эту идею, известную под названием ecological fitting (что можно приблизительно перевести как «экологическое соответствие» или «экологический подбор»), начиная с 1980-х годов развивает известный американский эколог Дэниел Джензен (Daniel Janzen).
Могут ли облигатно-мутуалистические системы, обычно считающиеся чем-то вроде апофеоза коэволюции, формироваться по такой же схеме, то есть без всякой коэволюции - просто за счет случайного соответствия двух случайно встретившихся видов, которые при определенных условиях оказываются неспособными жить друг без друга? Эксперименты, проведенные биологами из Гарвардского университета (США), позволяют ответить на этот вопрос утвердительно.
Авторы работали с обычными пекарскими почкующимися дрожжами Saccharomyces cerevisiae и не менее обычными одноклеточными водорослями хламидомонадами (Chlamydomonas reinhardtii). В природе эти виды в мутуалистических отношениях замечены не были. В лаборатории, однако, они вступили в неразрывную связь легко и быстро, без всякой эволюции или генетических модификаций. Для этого оказалось достаточно выращивать дрожжи и хламидомонады без доступа воздуха в среде, где глюкоза является единственным источником углерода, а нитрит калия - единственным источником азота.
Схема мутуалистических взаимоотношений дрожжей и хламидомонад довольно проста (рис. 1). Дрожжи питаются глюкозой и производят углекислый газ, необходимый хламидомонадам для фотосинтеза (использовать содержащуюся в среде глюкозу хламидомонады не умеют). Водоросли, со своей стороны, восстанавливают нитрит, переводя азот в доступную для дрожжей форму (аммоний). Таким образом, дрожжи обеспечивают хламидомонады углеродом, а хламидомонады снабжают дрожжи азотом. В таких условиях ни один из видов не может расти без другого. Это и есть облигатный мутуализм.
Авторы убедились, что мутуалистическая система благополучно растет в широком диапазоне концентраций глюкозы и нитрита, хотя в одиночку ни один из двух видов в этих условиях не выживает. Только при очень сильном снижении концентрации глюкозы или нитрита рост смешанной культуры прекращается.
Если раскупорить систему, то есть предоставить ей доступ к атмосферному CO2, получается сообщество, в котором только один из участников (дрожжи) не может жить без другого, тогда как второй участник (хламидомонады) уже не нуждается в первом для выживания. Впрочем, даже в этом случае хламидомонады лучше растут в присутствии дрожжей, чем без них (очевидно, дополнительный CO2, выделяемый дрожжами, идет им на пользу). Таким образом, система остается мутуалистической, хотя со стороны водорослей мутуализм уже не облигатный. Ни один из видов не вытесняет другой.
Если добавить в среду аммоний, получается обратная ситуация: теперь дрожжи могут жить без водорослей (и вообще не нуждаются в них), тогда как водоросли по-прежнему не могут жить без дрожжей. Это уже не мутуализм, а комменсализм (нахлебничество со стороны водорослей). В этом случае дрожжи, которые размножаются быстрее водорослей, заполняют всё жизненное пространство, доводя хламидомонады до вымирания. Авторы предполагают, что устойчивость таких асимметричных систем (в которых только один из участников сильно зависит от другого) определяется соотношением скоростей размножения. Если зависимый вид размножается быстрее, чем независимый, то сожительство двух видов может быть устойчивым; в противном случае независимый вид может полностью вытеснить своего напарника.
Авторы провели аналогичные эксперименты с другими видами хламидомонад и грибов-аскомицетов. Оказалось, что почти все виды дрожжей в данных условиях образуют облигатно-мутуалистические взаимоотношения с хламидомонадами. Правда, продуктивность (скорость роста) симбиотических комплексов оказывается разной. От чего она зависит, определить не удалось: авторы не нашли связи ни со склонностью дрожжей к кислородному дыханию или бескислородному метаболизму (брожению), ни с природными местообитаниями дрожжей, ни со скоростью размножения, ни со степенью влияния концентрации нитритов на рост дрожжей. Очевидно, дело в каких-то других особенностях изученных видов.
Одноклеточная водоросль хлорелла отказалась вступать в мутуалистические отношения с дрожжами, потому что она сама умеет питаться глюкозой и в смешанной культуре вытесняет дрожжи. Не стали образовывать облигатно-мутуалистические комплексы с водорослями дрожжи Hansenula polymorpha, потому что они сами умеют использовать нитрит в качестве источника азота. Но все же исследование показало, что самые разные виды аскомицетов и хламидомонад готовы вступить в симбиотические отношения друг с другом, попав в подходящие условия.
Из многоклеточных (точнее, образующих нитчатые гифы) аскомицетов были протестированы два классических лабораторных объекта - Neurospora crassa и Aspergillus nidulans. Оба вида умеют восстанавливать нитрит и потому не образуют облигатно-мутуалистических систем с хламидомонадами. Однако генетически модифицированные штаммы этих грибов, лишенные способности утилизировать нитрит, вступили в симбиоз с водорослями точно так же, как и дрожжи. Как выяснилось, при этом клетки хламидомонад вступают в непосредственный физический контакт с гифами грибов: под микроскопом видны гифы, обвешанные хламидомонадами, как новогодняя елка (рис. 2).
Мутуалистические взаимоотношения хламидомонад с дрожжами, по-видимому, тоже требуют установления физических контактов между клетками. Об этом свидетельствует тот факт, что систематическое встряхивание смешанной культуры дрожжей и водорослей резко замедляет рост симбиотической системы.
При помощи электронного микроскопа авторы обнаружили плотные контакты, образующиеся между клеточными стенками Aspergillus nidulans и Chlamydomonas reinhardtii, причем клеточная стенка водоросли в местах контакта становится тоньше - возможно, под действием ферментов, выделяемых грибом.
Похожие межклеточные контакты характерны для классических грибно-водорослевых симбиотических систем - лишайников. Аскомицеты в ходе своей эволюции много раз вступали в симбиоз с водорослями и цианобактериями, образуя лишайники. Лишайникообразующие группы разбросаны по всему филогенетическому дереву аскомицетов. Это значит, что такие эволюционные события происходили многократно и независимо в разных эволюционных линиях грибов (см. F. Lutzoni et al., 2001. Major fungal lineages are derived from lichen symbiotic ancestors). По-видимому, аскомицеты в целом «предрасположены» (преадаптированы) к формированию мутуалистических комплексов с одноклеточными водорослями. Эксперименты американских ученых, возможно, проливают свет на ранние стадии формирования таких комплексов.
Впрочем, не следует переоценивать сходство полученных в эксперименте мутуалистических систем с лишайниками. Хотя бы потому, что у большинства лишайников только грибной компонент не может жить в одиночку, тогда как фотосинтезирующие компоненты (одноклеточные водоросли и цианобактерии), как правило, могут прекрасно жить и без гриба. То есть лишайники не являются облигатно-мутуалистическими системами. Да и отсутствие доступа к атмосферному CO2 вряд ли является проблемой, с которой водорослям часто приходится сталкиваться в природе. Главное в обсуждаемой работе - демонстрация общего принципа. Исследование показало, что облигатный мутуализм может сложиться мгновенно, без всякой эволюции - просто за счет того, что изменившиеся условия делают виды взаимозависимыми. Разумеется, для того, чтобы из такого наспех сформированного симбиотического комплекса развилось что-то действительно сложное и высоко интегрированное, вроде лишайника, без миллионов лет коэволюции уже не обойтись.


Симбиоз - это длительное сожительство организмов двух или нескольких разных видов растений или животных, когда их отношения друг с другом очень тесны и обычно взаимно выгодны. Симбиоз обеспечивает этим организмам лучшее питание. Благодаря симбиозу организмам легче преодолевать неблагоприятные воздействия окружающей среды.
В тропических странах встречается очень интересное растение - мирмекодия. Это растение-муравейник. Живет оно на ветках или стволах других растений. Нижняя часть его стебля сильно расширена и представляет собой как бы большую луковицу. Вся луковица пронизана каналами, сообщающимися между собой. В них и поселяются муравьи. Эти каналы возникают в процессе развития утолщенного стебля, а не прогрызаются муравьями. Следовательно, муравьи получают от растения готовое жилище. Но и растению приносят пользу живущие в нем муравьи. Дело в том, что в тропиках водятся муравьи-листорезы. Они приносят большой вред растениям. В мирмекодии поселяются муравьи другого вида, враждующие с муравьями-листорезами. Постояльцы мирмекодии не допускают листорезов к ее вершине и не дают им объесть ее нежные листья. Таким образом, растение предоставляет животному помещение, а животное защищает растение от его врагов. Кроме мирмекодии в тропиках растет немало и других растений, находящихся в содружестве с муравьями.
Растение-муравейник - мирмекодия: 1 - два растения, поселившиеся на одной ветке дерева; 2 - разрез стебля мирмекодии.
Встречаются еще более тесные формы симбиоза растений и животных. Таков, например, симбиоз одноклеточных водорослей с амебами, солнечниками, инфузориями и другими простейшими животными. В этих одноклеточных животных поселяются зеленые водоросли, например зоохлорелла. Долгое время зеленые тельца в клетках простейших животных считались органоидами, т. е. постоянными частями самого одноклеточного животного, и лишь в 1871 г. известный русский ботаник Л. С. Ценковский установил, что здесь имеет место сожительство разных простейших организмов. Впоследствии это явление было названо симбиозом.
Зоохлорелла, живущая в теле простейшего животного амебы, лучше защищена от неблагоприятных внешних воздействий. Тело амебы прозрачно, поэтому процесс фотосинтеза протекает у водоросли нормально. Животное получает от водоросли растворимые продукты фотосинтеза (главным образом углеводы - сахар) и питается ими. Кроме того, при фотосинтезе водоросль выделяет кислород, и животное использует его для дыхания. В свою очередь животное обеспечивает водоросль необходимыми для ее питания азотистыми соединениями. Взаимная выгода для животного и растения от симбиоза очевидна.

Водоросли в теле животных: 1 - амеба, a - водоросль зоохлорелла, б - ядро амебы, в - сократительная вакуоля амебы; 2 - корненожка паулинелла, a - ядро корненожки, б - зеленые водоросли, в - псевдоподии корненожки.
К симбиозу с водорослями приспособились не только простейшие одноклеточные животные, но и некоторые многоклеточные. Водоросли встречаются в клетках гидр, губок, червей, иглокожих и моллюсков. Для некоторых животных симбиоз с водорослями стал настолько необходим, что их организм не может развиваться нормально, если в его клетках нет водорослей.

Вверху - симбиоз в жизни низших растений. Лишайники: 1 - кладония; 2 - пармелия; 3 - ксаятория; 4 - цепочки и шарообразные клетки водорослей, видимые в микроскоп в срезе слоевищ различных лишайников. Внизу - растения из семейства орхидей: 1 - эпифитные тропические орхидеи с воздушными (а) и лентовидными (б) корнями; 2 - наземная орхидея умеренного пояса - венерин башмачок.
Особенно интересен симбиоз, когда оба его участника - растения. Пожалуй, самый разительный пример симбиоза двух растительных организмов - это лишайник. Лишайник всеми воспринимается как единый организм. На самом же деле он состоит из гриба и водоросли. Основу его составляют переплетающиеся гифы (нити) гриба. На поверхности лишайника эти гифы переплетены плотно, а в рыхлом слое под поверхностью среди гиф гнездятся водоросли. Чаще всего это одноклеточные зеленые водоросли. Реже встречаются лишайники с многоклеточными синезелеными водорослями. Клетки водорослей оплетены гифами гриба. Иногда на гифах образуются присоски, которые проникают внутрь клеток водорослей. Сожительство выгодно и грибу и водоросли. Гриб дает водоросли воду с растворенными минеральными солями, а получает от водоросли органические соединения, вырабатываемые ею в процессе фотосинтеза, главным образом углеводы.
Симбиоз так хорошо помогает лишайникам в борьбе за существование, что они способны поселяться на песчаных почвах, на голых, бесплодных скалах, на стекле, на листовом железе, т. е. там, где никакое другое растение существовать не может. Встречаются лишайники на Крайнем Севере, в высоких горах, в пустынях - лишь бы был свет: без света водоросль в лишайнике не может усваивать углекислый газ и погибает. Гриб и водоросль так тесно сжились в лишайнике, настолько представляют собой единый организм, что даже и размножаются они чаще всего совместно.
Долгое время лишайники принимали за обычное растение и относили их к мхам. Зеленые клетки в лишайнике принимались за хлорофилловые зерна зеленого растения. Лишь в 1867 г. такой взгляд был поколеблен исследованиями русских ученых А. С. Фаминцына и О. В. Баранецкого. Им удалось выделить зеленые клетки из лишайника ксантории и установить, что они могут не только жить вне тела лишайника, но и размножаться делением и спорами. Следовательно, зеленые клетки лишайника - самостоятельные водоросли.
Каждый знает, например, что подосиновики нужно искать там, где растут осины, подберезовики - в березовых лесах. Оказывается, что шляпочные грибы растут вблизи определенных деревьев не случайно. Те «грибы», которые мы собираем в лесу, - только их плодовые тела. Само же тело гриба - грибница, или мицелий, - живет под землей и представляет собой нитевидные гифы, пронизывающие почву (см. ст. «Грибы»). От поверхности почвы они тянутся к кончикам древесных корней. Под микроскопом видно, как гифы, словно войлоком, оплетают кончик корня Симбиоз гриба с корнями высших растений называют микоризой (в переводе с греческого - «грибокорень»).
Подавляющее большинство деревьев в наших широтах и очень много травянистых растений (в том числе и пшеница) образуют с грибами микоризу. Ученые установили, что нормальный рост многих деревьев невозможен без участия гриба, хотя есть деревья, которые могут развиваться и без них, например береза, липа. Симбиоз гриба с высшим растением существовал еще на заре наземной флоры. Первые высшие растения - псилотовые - уже имели подземные органы, тесно связанные с гифами грибов. Чаще всего гриб лишь оплетает корень своими гифами и образует чехол, как бы наружную ткань корня. Реже встречаются формы симбиоза, когда гриб поселяется в самих клетках корня. Особенно ярко такой симбиоз выражен у орхидей, которые вообще не могут развиваться без участия гриба.
Можно предполагать, что гриб использует для своего питания углеводы (сахар), выделяемые корнями, а высшее растение получает от гриба продукты разложения азотистых органических веществ в почве. Сам древесный корень получить эти продукты не может. Предполагают также, что грибы вырабатывают витаминоподобные вещества, усиливающие рост высшего растения. Кроме того, несомненно, что грибной чехол, облекающий корень дерева и имеющий многочисленные разветвления в почве, намного увеличивает поверхность корневой системы, поглощающей воду, что очень существенно в жизни растения.
Симбиоз гриба и высшего растения следует учитывать во многих практических мероприятиях. Так, например, при разведении леса, при закладке полезащитных лесных полос обязательно надо «заразить» почву грибами, вступающими в симбиоз с той породой деревьев, которую сажают.
Огромное практическое значение имеет симбиоз усваивающих азот бактерий с высшими растениями из семейства бобовых (бобы, горох, фасоль, люцерна и многие другие). На корнях бобового растения обычно возникают утолщения - клубеньки, в клетках которых находятся бактерии, обогащающие растение, а затем и почву азотом (см. ст. «Как устроено и питается зеленое растение»).
За прошедшие со времен Борне 100 лет в слоевище лишайников было открыто и описано много различных форм абсорбционных, или всасывающих, гиф гриба. Эти гифы плотно прижимаются к клетке водоросли или проникают в нее и служат, как предполагают, для передачи веществ, которые образуют водоросли в результате своей жизнедеятельности, грибному компоненту.
Гриб должен использовать лишь часть водорослей, оставляя резерв - здоровые и нормальные водоросли, содержимым которых он мог бы питаться.
Учеными были замечены любопытные защитные реакции со стороны лишайниковых водорослей. Например, одновременно с проникновением гаустория в клетку водоросли эта клетка делилась. При этом плоскость деления, как правило, проходила как раз через участок, занятый гаусторием, а образовавшиеся в результате этого процесса дочерние клетки были свободны от гаусториев. Было замечено также, что обычно гриб поражает водоросли, уже достигшие определенной стадии зрелости. В молодых растущих водорослях происходит энергичное отложение веществ в оболочке клетки и быстрое ее утолщение. Эта толстая оболочка клетки фикобионта препятствует проникновению абсорбционных органов гриба. Однако большей частью защитная реакция водорослей против активности грибного компонента очень слаба.
Однако все высказанные по этому поводу точки зрения до сих пор остаются лишь догадками и большей частью не подтверждены экспериментально: лишайники оказались очень трудным объектом для физиологических исследований. Ученые пока не научились выращивать и поддерживать в живом состоянии слоевище лишайников в искусственных условиях. Тот контакт между грибом и водорослью, который с такой легкостью достигается в природе (достаточно вспомнить многообразие лишайников!), никак не удается воспроизвести в лабораторных условиях. Наоборот, при переносе лишайников в лабораторию этот контакт легко нарушается и растение просто погибает. Время от времени появляются сообщения об удачных опытах выращивания лишайника в условиях лаборатории, но пока эти сообщения единичны и не всегда достоверны.
Одной из причин неудач подобных попыток можно считать чрезвычайно медленный рост лишайников. Лишайники - многолетние растения. Обычно возраст взрослых слоевищ, которые можно увидеть где-нибудь в лесу на стволе деревьев или на почве, составляет не менее 20-50 лет. В северных тундрах возраст некоторых кустистых лишайников рода кладония достигает 300 лет. Слоевище лишайников, имеющих вид корочки, в год дает прирост всего 0,2-0,3 мм.
Кустистые и листоватые лишайники растут несколько быстрее - в год их слоевище увеличивается на 2-3 мм. Поэтому, чтобы вырастить взрослый лишайник в лаборатории, требуется не менее 20 лет, а может быть, и вся жизнь исследователя. Трудно проводить столь долговременный эксперимент!
Вот почему физиологические особенности лишайников, в том числе взаимоотношения компонентов, как правило, изучают на культурах изолированных мико- и фикобионтов. Этот метод очень перспективен, так как позволяет ставить длительные и воспроизводимые опыты. Но, к сожалению, данные, полученные этим методом, не могут полностью отразить те процессы, которые происходят в целом слоевище лишайника.
И тем более мы не вправе считать, что в природе, в естественных условиях, в слоевищах лишайника эти процессы протекают точно так же, как в культурах изолированных симбионтов. Вот почему все теории, пытающиеся объяснить взаимоотношения компонентов лишайников, остаются пока лишь догадками.
Более успешным оказалось изучение форм контакта между гифами гриба и клетками водорослей в слоевищах лишайников. Как показали исследования с применением электронной микроскопии, в слоевище лишайников можно встретить по крайней мере пять типов контакта между грибными гифами и водорослевыми клетками (рис. 289).
Чаще всего отдельная клетка водоросли и клетка грибной гифы находятся в непосредственном контакте друг с другом. В таком случае гриб образует специальные абсорбционные, всасывающие органы, которые проникают внутрь водорослевой клетки или плотно прижимаются к ее оболочке.
В настоящее время среди абсорбционных органов гриба в слоевище лишайников различают несколько типов: гаустории, импрессории и аппрессории.
Гаустории - это боковые выросты гиф гриба, которые прорывают оболочку клетки водоросли и проникают в ее протопласт (рис. 289, 2). Обычно в клетке водоросли развивается один гаусторий, но иногда их может быть и два. В слоевище лишайника гаустории встречаются в большом количестве и существуют продолжительное время. Было замечено, что в оболочках молодых гаусториев нет отложений целлюлозы, которая могла бы затруднять обмен между клеткой водоросли и гифой гриба. Старые гаустории почти всегда одеты довольно толстым слоем целлюлозы. Различают интрацеллюлярные (внутриклеточные) и интрамембранные (внутриоболочковые) гаустории.
У более высокоорганизованных лишайников образуются только интра мембранные гаустории. Они прорывают оболочку клетки водоросли и достигают ее протопласта, но не углубляются в него, а остаются в оболочке водорослевой клетки (рис. 289, 5). Наибольшее количество интрамембранных гаусториев образуется в слоевище лишайников весной, в начале вегетационного периода. С наступлением осени они далеко отступают от протопласта водоросли.
Другой тип всасывающих органов гриба - импрессорий - тоже боковые выросты грибных гиф, но, в отличие от гаусториев, они пе прорывают оболочку клетки водоросли, а вдавливают ее внутрь (рис. 289, 6, 7). Импрессорий отмечены у очень многих лишайников, например у широко распространенной пельтигеры (Peltigera).
Интересно, что в слоевищах, произрастающих во влажных местообитаниях, импрессории почти не развиваются, у тех же видов в сухих местообитаниях они образуются в большом количестве. При длительной засухе число импрессориев также увеличивается. Предполагают, что в засушливые периоды и в сухих местообитаниях гриб, чтобы удовлетворить потребности в питании, увеличивает свою всасывающую поверхность за счет увеличения количества и размеров импрессориев.
В отличие от гаусториев и импрессориев, образованных боковыми отростками гифы, аппрессории образуются вершиной грибной гифы. Такая вершина гифы плотно прижимается снаружи к оболочке клетки водоросли, никогда не проникая ни в ее протопласт, ни в ее внутренний слой (рис. 289, 8).
По в более старых участках слоевища можно найти немало отмерших обесцвеченных клеток - гриб рано или поздно все-таки убивает водоросли.
Такой же тип контакта между гифами гриба и клетками водорослей был найден у некоторых слизистых и базидиальных лишайников.
У ряда лишайников, в слоевище которых встречаются нитчатые улотриксовые водоросли, можно наблюдать еще один тип контакта. Как правило, в таком случае нити водорослей бывают целиком покрыты грибными гифами. Причем лишь иногда гифы образуют на поверхности водорослевой пити рыхлую сетку. Чаще же они располагаются очень густо и, срастаясь своими стенками, образуют сплошной чехол. Отдельная лопасть такого лишайника имеет вид тончайшего волоса. Под микроскопом она напоминает полую трубку, стенки которой образованы сросшимися грибными гифами; внутри трубки тянется нить водоросли.

У слизистых лишайников семейства коллемовых (Collemataceae) обычно не наблюдается никакого контакта между грибными гифами и клетками водорослей. Слоевище коллемовых не имеет дифференцированной структуры: нити водоросли посток разбросаны в беспорядке среди грибных гиф по всей толще слоевища (рис. 297, 2). Никаких абсорбционных отростков в клетках водорослей обычно пе заметно; гифы гриба и нити сине-зеленой водоросли расположены друг около друга, не вступая в видимый контакт. Предполагают, что в данном случае гриб поглощает органические вещества, ассимилируемые водорослями, прямо из слизи, которая обычно окружает нити ностока. Однако более тщательное изучение этих лишайников показало, что у многих видов коллемы (Collema) в слоевище время от времени образуются специальные абсорбционные гифы, которые тесно прижимаются к одной из клеток водорослевой пити, а через некоторое время можно наблюдать отмирание этой клетки.
Описанные выше формы контакта между гифами микобионта и клетками водорослей, по всей видимости, не исчерпывают всего многообразия способов, с помощью которых гриб и водоросль в слоевище лишайников устанавливают между собой тесную связь. Исследования в этом направлении только начинаются. Можно думать, что дальнейшее изучение тончайших структур лишайникового слоевища с помощью электронного микроскопа не только даст много нового в описании физических контактов между грибным и водорослевым компонентами лишайников, но и откроет новые горизонты в понимании их взаимоотношений.
Жизнь растений: в 6-ти томах. - М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров . 1974 .
Все составляющие части животного и растительного мира находятся в тесной взаимосвязи между собой и вступают в сложные взаимоотношения. Некоторые благоприятны для участников или вообще жизненно важны, например лишайники (представляют собой результат симбиоза гриба и водоросли), другие безразличны, третьи же приносят вред. Исходя из этого, принято различать три вида взаимоотношений организмов - это нейтрализм, антибиоз и симбиоз. Первый, по сути, не представляет ничего особенного. Это такие отношения между популяциями, обитающими на одной территории, при которых они не влияют друг на друга, не взаимодействуют. А вот антибиоз и симбиоз - примеры, которых встречаются очень часто, являются важными компонентами естественного отбора и участвуют в дивергенции видов. Остановимся на них более подробно.
Симбиоз: что это такое?
Представляет собой достаточно распространенную форму взаимовыгодного сожительства организмов, при которой существование одного партнера невозможно без другого. Наиболее известный случай - это симбиоз гриба и водоросли (лишайники). Причем первый получает продукты фотосинтеза, синтезируемые вторым. А водоросль извлекает минеральные соли и воду из гиф гриба. Жизнь по отдельности невозможна.
Комменсализм
Комменсализм - это фактически одностороннее использование одним видом другого, без оказания на него вредного воздействия. Может осуществляться в нескольких формах, но основных две:

Все остальные в какой-то мере являются модификациями этих двух форм. Например, энтойкия, при которой один вид обитает в теле другого. Наблюдается это у рыбок карапус, которые используют в качестве жилища клоаку голотурий (вид иглокожих), но питаются за ее пределами различными мелкими рачками. Или эпибиоз (одни виды живут на поверхности у других). В частности, усоногие рачки хорошо себя чувствуют на горбатых китах, абсолютно им не мешая.
Кооперация: описание и примеры
Кооперация - это такая форма взаимоотношений, при которой организмы могут прожить отдельно, но иногда объединяются для общей пользы. Получается, что это необязательный симбиоз. Примеры:
Взаимное сотрудничество и совместное проживание в животной среде не редкость. Приведем лишь некоторые наиболее интересные примеры.

Симбиотические отношения между растениями
Симбиоз растений очень распространен, и если приглядеться внимательно к окружающему нас миру, то можно невооруженным глазом увидеть его.

Симбиоз (примеры) животных и растений

Примеры очень многочисленны, и многие отношения между разными элементами растительного и животного мира еще мало изучены.
Что такое антибиоз?

Симбиоз, примеры которого встречаются практически на каждом шагу, в том числе и в жизни человека, в составе естественного отбора является важным компонентом эволюции в целом.
Все живые организмы на планете поделены на царства. В основу классификации было взято наличие ядра. Есть царство прокариотов, не имеющих ядра. К ним относят бактерии и сине-зеленые водоросли (цианеи). К царству эукариотов относятся те организмы, которые имеют ядро: грибы, растения и животные. Несмотря на то что бактерии, грибы, растения (водоросли и высшие), животные составляют отдельные царства, между ними есть и общие черты.
Бактерии и цианеи относят к прокариотам. Их основными отличиями являются:
- отсутствие четко оформленного ядра;
- отсутствие мембранных органоидов;
- наличие мезосом (своеобразные выпячивания мембраны в середину клетки);
- мелкие рибосомы, по сравнению с эукариотами;
- у бактерий одна хромосома, у цианобактерий несколько хромосом, которые находятся в цитоплазме;
- отсутствие ядрышек;
- нет митохондрий;
- клеточная стенка у бактерий состоит из муреина, а у цианей из целлюлозы;
- жгутики отличаются простотой строения и малым диаметром;
- полового процесса нет, размножение происходит с помощью деления.

При неблагоприятных условиях многие микроорганизмы образуют споры, которые могут лежать годами в ожидании подходящих условий для жизни и развития. Растения и грибы также образуют споры, но они им необходимы для размножения. Есть микробы, которые питаются так же, как и растения, и являются автотрофами, а некоторые питаются как животные и являются гетеротрофами. В отличие от других живых организмов, чья жизнь без наличия кислорода невозможна, существуют микроорганизмы, которые способны проживать в анаэробной среде, и кислород, наоборот, для них губителен.
Бактерии являются самыми многочисленными существами на планете, и большинство из них до сих пор не изучено.
Царство растений
В основу классификации положено их основное отличие ─ это автотрофное питание. Они способны перерабатывать неорганические вещества в органические. Для этого им необходима солнечная энергия. Это свойственно и цианобактериям. Благодаря растениям и цианобактериям воздух на планете обогащается кислородом, который так необходим другим живым организмам. Растения являются источником пищи для многих других организмов. Их разделяют на два подцарства: водоросли и высшие. Водоросли не имеют корня, стебля и листьев в отличие от высших форм.

Отдельное место занимают примитивные водоросли (пиррофитовые), у которых в клетках в хромосомах отсутствуют гистоны, по своей структуре они близки к нуклеоиду бактерий. Клеточная стенка некоторых водорослей состоит из хитина, как у животных и грибов. Красные водоросли отличаются от остальных видов тем, что их клетки не имеют жгутиков. Имеются отличия в особенностях строения и биохимических процессах.
Царство грибов
Долгое время ученые спорили по поводу того, выделять грибы в отдельное царство или нет. В результате долгих споров их все же выделили отдельно, так как они имеют много общего и с растениями, и с животными.

Способ питания у них такой же, как и у животных ─ гетеротрофный. Так же, как и у животных, у них отсутствуют пластиды и в клеточной стенке есть хитин. В результате обменных процессов образуется мочевина. Грибы, так же как и растения, поглощают питательные вещества путем всасывания. Они неподвижны и имеют способ роста такой же, как и у растений.
Некоторые грибы размножаются как бактерии ─ бесполо, некоторые как растения ─ вегетативным путем, некоторые как животные ─ половым. Многие из них, как и микробы, перерабатывают отмершие живые организмы, тем самым играют роль «санитаров». Многие из них приносят пользу и применяются в производстве антибиотиков, гормонов, витаминов.
В зависимости от того, как они потребляют органические вещества, их делят на три вида:
Лишайники
Многие ученые настаивают на том, чтобы выделить лишайники в отдельное царство. На это есть несколько причин. Они могут быть симбионтами:
- гриба и водоросли;
- бактерий гриба и водорослей.
По внешнему виду их делят на три группы:
- корковые (которые растут на камнях и прочно срастаются с поверхностью);
- листоватые (прикрепляются к поверхности с помощью ножки);
- кустистые (прикрепляются к почве, деревьям, кустарникам в виде кустиков).

Тело лишайника называется слоевищем, которое у разных видов отличается по размерам, окраске, форме и по строению. Слоевище может быть от нескольких сантиметров до метра.
Растут лишайники очень медленно, но длительность жизни может быть от сотни до тысячи лет.
В результате симбиоза получается единый организм. Причем гифы гриба тесно переплетаются с клетками водорослей. Таким образом, в лишайнике сочетаются два совершенно разных организма по строению и способу питания. Грибы, составляющие симбиоз с водорослями, отдельно в природе не встречаются, а вот виды водорослей, участвующих в симбиозе, можно встретить и как отдельно живущий организм.
У лишайников уникальный способ питания: грибы всасывают растворенные минералы, а цианобактерии образуют органику и участвуют в процессе фотосинтеза. Размножаться лишайники могут как спорами, так и делением слоевища.

Чувствительность лишайников к загрязненной окружающей среде делает их индикаторами чистоты. Многие виды используют для питания животных и в медицинских целях.
Царство животных
Царство животных подразделяют на два подцарства: простейшие и многоклеточные. Несмотря на то что простейшие состоят из одной клетки, так же как и бактерии, они имеют все характеристики, свойственные животным. Есть виды простейших, которые на свету питаются автотрофно, а при его отсутствии переходят на гетеротрофное. Размножаться простейшие могут как бесполым способом ─ делением клетки, так и половым ─ конъюгация.

Общее у животных и растений ─ это обмен веществ и строение клеток. Основное отличие ─ это способ питания. Животные – гетеротрофы, то есть они питаются уже готовыми органическими соединениями и не способны синтезировать неорганические вещества. В большинстве своем они подвижны.
Более сложное строение клетки эукариотов говорит о том, что эти усовершенствования они получали в результате эволюции. А одновременное существование на земле как прокариот, так и эукариот говорит о том, что биологические процессы свойственны всем формам жизни. Все живые организмы живут в полном взаимодействии друг с другом, и исчезновение хотя бы одного из видов привело бы к необратимым последствиям. На планете есть место всем видам экологической цепочки.