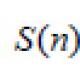Спинной мозг морфофункциональная организация спинного мозга. Цитология как наука и ее взаимосвязь с другими науками
Спинной мозг состоит из двух симметричных половин, отграниченных друг от друга спереди - глубокой срединной щелью, а сзади - срединной бороздой. Спинной мозг характеризуется сегментарным (метамерным) строением (31-33 сегмента); с каждым сегментом связана пара передних (вентральных) и пара задних (дорсальных) корешков.
В спинном мозге различают серое вещество , расположенное в центральной части, и белое вещество , лежащее по периферии.
Внешнюю границу белого вещества спинного мозга образует пограничная глиальная мембрана , состоящая из слившихся уплощенных отростков астроцитов. Эту мембрану пронизывают нервные волокна, составляющие передние и задние корешки.
На протяжении всего спинного мозга в центре серого вещества проходит центральный канал спинного мозга, сообщающийся с желудочками головного мозга.
Серое вещество на поперечном разрезе имеет вид бабочки и включает передние , или вентральные, задние , или дорсальные, и боковые , или латеральные, рога. В сером веществе находятся тела, дендриты и (частично) аксоны нейронов, а также глиальные клетки. Основной составной частью серого вещества, отличающей его от белого, являются мультиполярные нейроны. Между телами нейронов находится нейропиль - сеть, образованная нервными волокнами и отростками глиальных клеток.
Среди всех нейронов спинного мозга можно выделить три вида клеток:
· корешковые,
· внутренние,
· пучковые.
Аксоны корешковых клеток покидают спинной мозг в составе его передних корешков, это клетки боковых и передних рогов. Отростки внутренних клеток заканчиваются синапсами в пределах серого вещества спинного мозга (преимущественно нейроны задних рогов). Аксоны пучковых клеток проходят в белом веществе обособленными пучками волокон, несущими нервные импульсы от определенных ядер спинного мозга в его другие сегменты или в соответствующие отделы головного мозга, образуя проводящие пути.
В процессе развития спинного мозга из нервной трубки изогенетически нейроны группируются в 10 слоях, или пластинах Рекседа . При этом I-V пластины соответствуют задним рогам, VI-VII пластины - промежуточной зоне, VIII-IX пластины - передним рогам, X пластина - зона около центрального канала. На поперечных срезах более отчетливо видны ядерные группы нейронов, а на сагиттальных - лучше видно пластинчатое строение, где нейроны группируются в колонки Рекседа.
Клетки, сходные по размерам, строению и функциональному значению, лежат в сером веществе группами, которые называются ядрами .
В задних рогах различают губчатый слой, желатинозное вещество, собственное ядро заднего рога и грудное ядро Кларка, ядро Роланда с тормозными нейронами, зону Лиссаауэра .
Нейроны губчатой зоны и желатинозного вещества осуществляют связь между чувствительными клетками спинальных ганглиев и двигательными клетками передних рогов, замыкая местные рефлекторные дуги.
Нейроны ядра Кларка получают информацию от рецепторов мышц, сухожилий и суставов (проприоцептивная чувствительность) по самым толстым корешковым волокнам и передают ее в мозжечок, это крупные мультиполярные нейроны.
Нейроны собственного ядра заднего рога это вставочные мелкие мультиполярные клетки, аксоны которых заканчиваются в пределах серого вещества спинного мозга той же стороны (ассоциативные клетки) или противоположной стороны (комиссуральные клетки).
Между задними и боковыми рогами серое вещество вдается тяжами в белое, вследствие чего образуется его сетеобразное разрыхление, получившее название сетчатого образования, или ретикулярной формации спинного мозга.
В промежуточной зоне (боковых рогах) расположены центры вегетативной (автономной) нервной системы - преганглионарные холинергические нейроны ее симпатического и парасимпатического отделов.
В передних рогах расположены самые крупные нейроны спинного мозга. Это корешковые клетки, поскольку их аксоны составляют основную массу волокон передних корешков. В передних рогах находятся 3 типа нейронов, формирующих значительные по объему 5 групп ядер (латеральные – передняя и задняя группа, медиальные – передняя и задняя группа и центральное или промежуточное ядро).
Альфа мотонейроны - крупные нейроны 100-140 мкм. По функции они двигательные и их аксоны в составе передних корешков выходят из спинного мозга и направляются к поперечно полосатым мышцам.
Гамма мотонейроны – более мелкие, являются клетками контролирующими силу и скорость сокращения.
клетки Реншоу - тормозные клетки осуществляют взаимное торможение мотонейронов сгибателей и разгибателей, а так же осуществляют возвратное торможение.
Белое вещество рогами мозга разделено на столбы: передние (нисходящие), средние (смешанные) и задние (восходящие). Белое вещество спинного мозга представляет собой совокупность продольно ориентированных преимущественно миелиновых нервных волокон. Пучки нервных волокон, осуществляющие связь между различными отделами нервной системы, называются трактами, или проводящими путями, спинного мозга.
4. Рефлекторный аппарат спинного мозга (соматические рефлекторные дуги )
Элементарная рефлекторная дуга собственного аппарата спинного мозга представлена двумя нейронами. Тело первого – афферентного нейрона расположено в спинальном ганглии. Его дендрит направляется на периферию и заканчивается рецептором. Аксон афферентного нейрона в составе задних корешков входит в спинной мозг, его задние рога, и проходит транзитом до клеток передних рогов спинного мозга. В передних рогах расположены тела двигательных эфферентных клеток – крупных альфа-мотонейронов, на которых и заканчивается аксосоматическим синапсом аксон чувствительной клетки. Аксон эфферентного нейрона покидает спинной мозг, входит в состав передних корешков, далее в спинномозговой нерв, сплетение и, наконец, в составе соматического нерва достигает органа эффектора (мышцы, железы).
При нанесении раздражения (укол пальца кисти) происходит раздражение рецепторного аппарата (ноцерецепторов кожи) и генерация нервного импульса, который центростремительно через дендрит, тело афферентного нейрона и его аксон проводится посредством синаптической связи до тела второго эфферентного нейрона. Оттуда нервный импульс центробежно посредством аксона клетки покидает спинной мозг, передний корешок, нерв и вызывает возбуждение в органе эффекторе (двуглавой мышце плеча), что, в свою очередь, приводит к ожидаемому эффекту – отдергиванию руки.
Принцип строения и работы вегетативных рефлекторных дуг разбирается самостоятельно.
Спинной мозг - наиболее древнее образование центральной нервной системы; он впервые появляется у ланцетника
Характерной чертой организации спинного мозга является периодичность его структуры в форме сегментов, имеющих входы в виде задних корешков, клеточную массу нейронов (серое вещество) и выходы в виде передних корешков.
Спинной мозг человека имеет 31-33 сегмента: 8 шейных, 12 грудных, 5 поясничных,5 крестцовых 1-3 копчиковых.
Морфологических границ между сегментами спинного мозга не существует. Каждый сегмент через свои корешки иннервирует три метамера тела и получает информацию также от трех метамеров тела. В итоге каждый метамер тела иннервируется тремя сегментами и передает сигналы в три сегмента спинного мозга.
Задние корешки являются афферентными, чувствительными, центростремительными, а передние - эфферентными, двигательными, центробежными (закон Белла-Мажанди).
Афферентные входы в спинной мозг организованы аксонами спинальных ганглиев, лежащих вне спинного мозга, и аксонами симпатического и парасимпатического отделов вегетативной нервной системы.
Первая группа афферентных входов спинного мозга образована чувствительными волокнами, идущими от мышечных рецепторов, рецепторов сухожилий, надкостницы, оболочек суставов. Эта группа рецепторов образует начало так называемой проприоцептивной чувствительности.
Вторая группа афферентных входов спинного мозга начинается от кожных рецепторов: болевых, температурных, тактильных, давления.
Третья группа афферентных входов спинного мозга представлена волокнами от висцеральных органов, это висцеро-рецептивная система.
Эфферентные (двигательные) нейроны расположены в передних рогах спинного мозга, и их волокна иннервируют всю скелетную мускулатуру.
Особенности нейронной организации спинного мозга
Нейроны спинного мозга образуют его серое вещество в виде симметрично расположенных двух передних и двух задних рогов. ядра, вытянутые по длине спинного мозга, и на поперечном разрезе располагается в форме буквы Н. В грудном отделе спинной мозг имеет, помимо названных, еще и боковые рога.
Задние рога выполняют главным образом сенсорные функции, от них передаются сигналы в вышележащие центры, в структуры противоположной стороны, либо к передним рогам спинного мозга.
В передних рогах находятся нейроны, дающие свои аксоны к мышцам. Все нисходящие пути центральной нервной системы, вызывающие двигательные реакции, заканчиваются на нейронах передних рогов. В связи с этим Шеррингтон назвал их «общим конечным путем».
В боковых рогах, начиная с I грудного сегмента спинного мозга и до первых поясничных сегментов, располагаются нейроны симпатического, а в крестцовых - парасимпатического отдела вегетативной нервной системы.
Спинной мозг человека содержит около 13 млн. нейронов, из них 3% - мотонейроны, а 97% - вставочные. Функционально нейроны спинного мозга можно разделить на 4 основные группы:
1) мотонейроны, или двигательные, - клетки передних рогов, аксоны которых образуют передние корешки;
2) интернейроны - нейроны, получающие информацию от спинальных ганглиев и располагающиеся в задних рогах. Эти нейроны реагируют на болевые, температурные, тактильные, вибрационные, проприоцептивные раздражения;
3) симпатические, парасимпатические нейроны расположены преимущественно в боковых рогах. Аксоны этих нейронов выходят из спинного мозга в составе передних корешков;
4) ассоциативные клетки - нейроны собственного аппарата спинного мозга, устанавливающие связи внутри и между сегментами.
В средней зоне серого вещества (между задним и передним рогами) спинного мозга имеется промежуточное ядро (ядро Кахаля) с клетками, аксоны которых идут вверх или вниз на 1-2 сегмента и дают коллатерали на нейроны ипси- и контралатеральной стороны, образуя сеть. Подобная сеть имеется и на верхушке заднего рога спинного мозга - эта сеть образует так называемое студенистое вещество (желатинозная субстанция Роланда) и выполняет функции ретикулярной формации спинного мозга.Средняя часть серого вещества спинного мозга содержит преимущественно короткоаксонные веретенообразные клетки они выполняют связующую функцию между симметричными отделами сегмента, между клетками его передних и задних рогов.
Мотонейроны. Аксон мотонейрона своими терминалями иннервирует сотни мышечных волокон, образуя мотонейронную единицу. Несколько мотонейронов могут иннервировать одну мышцу, в этом случае они образуют так называемый мотонейронный пул. Возбудимость мотонейронов различна, поэтому при разной интенсивности раздражения в сокращение вовлекается разное количество волокон одной мышцы. При оптимальной силе раздражения сокращаются все волокна данной мышцы; в этом случае развивается максимальное сокращение. Мотонейроны могут генерировать импульсы с частотой до 200 в секунду.
Интернейроны. Эти промежуточные нейроны, генерирующие импульсы с частотой до 1000 в секунду, являются фоновоактивными и имеют на своих дендритах до 500 синапсов. Функция интернейронов заключается в организации связей между структурами спинного мозга и обеспечении влияния восходящих и нисходящих путей на клетки отдельных сегментов спинного мозга. Очень важной функцией интернейронов является торможение активности нейронов, что обеспечивает сохранение направленности пути возбуждения. Возбуждение интернейронов, связанных с моторными клетками, оказывает тормозящее влияние на мышцы-антагонисты.
Нейроны симпатического отдела вегетативной нервной системы расположены в боковых рогах грудного отдела спинного мозга, имеют редкую частоту импульсации (3-5 в секунду), парасимпатические нейроны локализуются в сакральном отделе спинного мозга.
При раздражении или поражениях задних корешков наблюдаются опоясывающие боли на уровне метамера пораженного сегмента, снижается чувствительность, исчезают или ослабляются рефлексы. Если происходит изолированное поражение заднего рога, утрачивается болевая и температурная чувствительность на стороне повреждения, а тактильная и проприоцептивная сохраняются, так как из заднего корешка аксоны температурной и болевой чувствительности идут в задний рог, а аксоны тактильной и проприоцептивной - прямо в задний столб и по проводящим путям поднимаются вверх.
Поражение переднего рога и переднего корешка спинного мозга приводит к параличу мышц, которые теряют тонус, атрофируются, при этом исчезают рефлексы, связанные с пораженным сегментом.
Поражение боковых рогов спинного мозга сопровождается исчезновением кожных сосудистых рефлексов, нарушением потоотделения, трофическими изменениями кожи, ногтей. Двустороннее поражение парасимпатического отдела на уровне крестцов приводит к нарушению дефекации и мочеиспускания.
Представляет собой уплощенный тяж , расположенный в спинномозговом канале, длиной около 45 см у мужчин и 42 см у женщин. В местах выхода нервов к верхним и нижним конечностям спинной мозг имеет два утолщения: шейное и поясничное.
Спинной мозг состоит из двух типов ткани : наружного белого (пучки нервных волокон) и внутреннего серого вещества (тела нервных клеток, дендриты и синапсы). В центре серого вещества вдоль всего мозга проходит узкий канал с цереброспинальной жидкостью. Спинной мозг имеет сегментарное строение (31-33 сегмента), каждый его участок связан с определенной частью тела, от сегментов спинного мозга отходит 31 пара спинномозговых нервов: 8 пар шейных (Ci-Cviii), 12 пар грудных (Thi-Thxii), 5 пар поясничных (Li-Lv), 5 пар крестцовых (Si-Sv) и пара копчиковых (Coi-Coiii).
Каждый нерв при выходе из мозга делится на передние и задние корешки . Задние корешки – афферентные пути, передние корешки эфферентные пути. По задним корешкам спинномозговых нервов в спинной мозг поступают афферентные импульсы от кожи, двигательного аппарата, внутренних органов. Передние корешки образованы двигательными нервными волокнами и передают эфферентные импульсы на рабочие органы. Чувствительные нервы преобладают над двигательными, поэтому происходит первичный анализ поступающих афферентных сигналов и формирование реакций наиболее важных для организма в данный момент (передача многочисленных афферентных импульсов на ограниченное число эфферентных нейронов называется конвергенция ).
Общее количество нейронов спинного мозга составляет около 13 млн. Их подразделяют: 1) по отделу нервной системы – нейроны соматической и вегетативной НС; 2) по назначению – эфферентные, афферентные, вставочные; 3) по влиянию – возбуждающие и тормозные.
Функции нейронов спинного мозга.
Эфферентные нейроны относятся к соматической нервной системе и иннервируют скелетные мышцы – мотонейроны. Различают альфа и гамма – мотонейроны. А-мотонейроны осуществляют передачу скелетным мышцам сигналов из спинного мозга. Аксоны каждого мотонейрона многократно делятся, поэтому каждый из них охватывает множество мышечных волокон, образуя с ним двигательную моторную единицу. Г-мотонейроны иннервируют мышечные волокна мышечного веретена. Они обладают высокой частотой импульсации, получают информацию о сотоянии мышечного веретена через промежуточные нейроны (вставочные). Генерируют импульсы с частотой до 1000 в сек. Это фоноактивные нейроны, имеющие на своих дендритах до 500 синапсов.
Афферентные нейроны соматической НС локализуются в спинальных ганглиях и ганглиях черепно-мозговых нервов. Их отростки проводят импульсацию от мышечных, сухожильных, кожных рецепторов, вступают в соответствующие сегменты спинного мозга и соединяются синапсами с вставочными или альфа-мотонейронами.
Функция вставочных нейронов состоит в организации связи между структурами спинного мозга.
Нейроны вегетативной нервной ситемы являются вставочными. Симпатические нейроны расположены в боковых рогах грудного отдела спинного мозга, они имеют редкую частоту импульсации. Одни из них участвуют в поддержании сосудистого тонуса, другие в регуляции гладкой мускулатуры пищеварительной системы.
Совокупность нейронов образует нервные центры.
В спинном мозге находятся центры регуляции большинства внутренних органов и скелетной мускулатуры. Центры управления скелетной мускулатурой находятся во всех отделах спинного мозга и иннервируют по сегментарному принципу скелетную мускулатуру шеи (Сi-Сiv), диафрагмы (Ciii-Cv), верхних конечностей (Cv-Thii), туловища (Thiii-Li), нижних конечностей (Lii-Sv). При повреждении определенных сегментов спинного мозга или его проводящих путей развиваются специфические двигательные нарушения и расстройства чувствительности.
Функции спинного мозга:
А) обеспечивает двустороннюю связь между спинномозговыми нервами и головным мозгом – проводниковая функция;
Б) осуществляет сложные двигательные и вегетативные рефлексы – рефлекторная функция.
Морфофункциональная организация спинного мозга
Спинной мозг – наиболее древний отдел ЦНС позвоночных. Он уже есть у ланцетника, наиболее примитивного представителя хордовых.
Спинной мозг – каудальный отдел ЦНС. Он помещается в позвоночном канале и имеет неодинаковую длину у разных представителей позвоночных.
У человека в каудальном отделе позвоночного канала собираются корешки каудальных отделов спинного мозга, образуя так называемый конский хвост.
Спинной мозг характеризуется сегментарным строением. В спинном мозге выделяют шейный, грудной, поясничный, крестцовый и копчиковый отделы. Каждый отдел состоит из нескольких сегментов. Шейный отдел включает 8 сегментов (С 1 – С 8), грудной – 12 (Th 1 – Th 12), поясничный – 5 (L 1 – L 5), крестцовый – 5 (S 1 – S 5) и копчиковый – 1-3 (Со 1 – Co 3). От каждого сегмента отходят две пары корешков, которые соответствуют одному из позвонков и покидают позвоночный канал через отверстие между ними.
Различают дорсальные (задние) и вентральные (передние) корешки. Дорсальные корешки образованы центральными аксонами первичных афферентных нейронов, тела которых лежат в спинномозговых ганглиях.
Вентральные корешки сформированы аксонами α- и γ-мотонейронов и немиелинизированными волокнами нейронов вегетативной нервной системы. Такое распределение афферентных и эфферентных волокон было установлено независимо друг от друга в начале XIX века Ч. Беллом (1811) и Ф. Мажанди (1822). Различное распределение функций в передних и задних корешках спинного мозга получило название закона Белла-Мажанди. Сегменты спинного мозга и позвонки соответствуют одному метамеру. Нервные волокна пары задних корешков идут не только к своему метамеру, но также выше и ниже – в соседние метамеры. Кожная область, в которой распределяются эти чувствительные волокна, называется дерматомом.
Количество волокон в дорсальном корешке гораздо больше, чем в вентральном.
Нейронные структуры спинного мозга. Центральную часть поперечного среза спинного мозга занимает серое вещество. Вокруг серого вещества располагается белое вещество. В сером веществе выделяют передние, задние и боковые рога, а в белом – столбы (вентральные, дорсальные, латеральные и т.д.).
Нейрональный состав спинного мозга достаточно разнообразен. Выделяют несколько видов нейронов. Тела нейронов спинномозговых ганглиев находятся вне спинного мозга. В спинной мозг входят аксоны этих нейронов. Нейроны спинномозговых ганглиев – это униполярные или псевдоуниполярные нейроны. В спинномозговых ганглиях лежат тела соматических афферентов, иннервирующих в основном скелетные мышцы. Тела других чувствительных нейронов находятся в ткани и в интрамуральных ганглиях автономной нервной системы и обеспечивают чувствительность только внутренних органов. Они бывают двух типов: крупные – диаметром 60-120 мкм и мелкие – диаметром 14-30 мкм. Крупные дают миелинизированные волокна, а мелкие – миелинизированные и немиелинизированные. Нервные волокна чувствительных клеток по скорости проведения и диаметру классифицируются на А-, В- и С-волокна. Толстые миелинизированные А-волокна диаметром от 3 до 22 мкм и скоростью проведения от 12 до 120 м/с подразделяются на подгруппы: альфа-волокна – от мышечных рецепторов, бета-волокна – от тактильных и барорецепторов, дельта-волокна – от терморецепторов, механорецепторов и болевых рецепторов. К волокнам группы В относят миелинизированные волокна средней толщины со скоростью проведения возбуждения 3-14 м/с. По ним в основном передается ощущение боли. К афферентам типа С относят большинство безмиелиновых волокон толщиной не более 2 мкм и скоростью проведения до 2 м/с. Это волокна, которые идут от болевых, хемо- и некоторых механорецепторов.
В сером веществе спинного мозга выделяют следующие элементы:
1) эфферентные нейроны (мотонейроны);
2) вставочные нейроны;
3) нейроны восходящих трактов;
4) интраспинальные волокна чувствительных афферентных нейронов.
Моторные нейроны сосредоточены в передних рогах, где они образуют специфические ядра, все клетки которых посылают свои аксоны к определенной мышце. Каждое двигательное ядро тянется обычно на протяжении нескольких сегментов, поэтому и их аксоны, которые иннервируют одну и ту же мышцу, покидают спинной мозг в составе нескольких вентральных корешков.
В промежуточной зоне серого вещества локализуются вставочные нейроны. Их аксоны распространяются как внутри сегмента, так и в ближайшие соседние сегменты. Вставочные нейроны – разнородная группа, дендриты и аксоны которой не покидают пределов спинного мозга. Вставочные нейроны образуют синаптические контакты только с другими нейронами, и их большинство. На долю вставочных нейронов приходится около 97% от всех нейронов. По размерам они меньше α-мотонейронов, способны к высокочастотной импульсации (выше 1000 в сек.). Для проприоспинальных вставочных нейронов характерно свойство посылать длинные аксоны через несколько сегментов и оканчиваться на мотонейронах. Вместе с тем на эти клетки конвергируют волокна различных нисходящих трактов. Поэтому они являются релейными станциями на пути от вышележащих нейронов к мотонейронам. Особую группу вставочных нейронов образуют тормозные нейроны. К ним относятся, например, клетки Реншоу.
Нейроны восходящих трактов также целиком находятся в пределах ЦНС. Тела этих нейронов расположены в сером веществе спинного мозга.
Центральные окончания первичных афферентов имеют свои особенности. После вступления в спинной мозг афферентное волокно, как правило, дает начало восходящей и нисходящей ветвям, которые могут идти на значительные расстояния по спинному мозгу. Концевые разветвления одного нервного афферентного волокна имеют многочисленные синапсы на одном мотонейроне. Кроме того, установлено, что одно волокно, идущее от рецептора растяжения, образует синапсы почти со всеми мотонейронами данной мышцы.
В дорсальной части дорсального рога располагается желатинозная субстанция Роланда.
Наиболее точное представление о топографии нервных клеток серого вещества спинного мозга дает разделение его на последовательные слои или пластины, в каждой из которых группируются, как правило, однотипные нейроны.
Согласно этим данным, все серое вещество спинного мозга было разделено на 10 пластин (Рексед) (рис. 2.2).
I – краевые нейроны – дают начало спиноталамическому тракту;
II-III – желатинозная субстанция;
I-IV – в целом первичная сенсорная область спинного мозга (афферентация от экстерорецепторов, афферентация от рецепторов кожной и болевой чувствительности);
Рис. 2.2. Деление серого вещества спинного мозга на пластины (по Рекседу)
V-VI – локализуются вставочные нейроны, которые получают входы от задних корешков и нисходящих путей (кортико-спинальный, рубро-спинальный);
VII-VIII – располагаются проприоспинальные вставочные нейроны (от проприорецепторов, волокон вестибуло-спинального и ретикуло-спи-
нального трактов), аксоны проприоспинальных нейронов;
IX – содержит тела α- и γ-мотонейронов, пресинаптические волокна первичных афферентов от мышечных рецепторов растяжения, окончания волокон нисходящих трактов;
X – окружает спинномозговой канал и содержит наряду с нейронами значительное количество глиальных клеток и комиссуральных волокон.
Свойства нервных элементов спинного мозга. Спинной мозг человека содержит примерно 13 миллионов нейронов.
α-мотонейроны – крупные клетки с длинными дендритами, имеющие до 20 000 синапсов, большая часть которых образована окончаниями интраспинальных вставочных нейронов. Скорость проведения по их аксону составляет 70-120 м/с. Характерны ритмические разряды с частотой не выше 10-20 имп/с, что связано с выраженной следовой гиперполяризацией. Это выходные нейроны. Они осуществляют передачу сигналов скелетным мышечным волокнам, выработанных в спинном мозге.
γ-мотонейроны – более мелкие клетки. Диаметр их не более 30-40 мкм, они не имеют непосредственного контакта с первичными афферентами.
γ-мотонейроны иннервируют интрафузальные (внутриверетенные) мышечные волокна.
Они моносинаптически активируются волокнами нисходящих трактов, что играет важную роль в α-, γ-взаимодействии. Скорость проведения по их аксону ниже – 10-40 м/с. Частота импульсов выше, чем у α-мото-
нейронов, – 300-500 имп/с.
В боковых и передних рогах находятся преганглионарные нейроны вегетативной нервной системы – аксоны их направляются к клеткам ганглиев симпатической нервной цепочки и к интрамуральным ганглиям внутренних органов.
Тела симпатических нейронов, аксоны которых образуют преганглионарные волокна, располагаются в интермедиолатеральном ядре спинного мозга. Их аксоны относятся к группе В-волокон. Им свойственна постоянная тоническая импульсация. Одни из этих волокон участвуют в поддержании сосудистого тонуса, а другие обеспечивают регуляцию висцеральных эффекторных структур (гладкой мускулатуры пищеварительной системы, железистых клеток).
Тела парасимпатических нейронов образуют крестцовые парасимпатические ядра. Они располагаются в сером веществе крестцового отдела спинного мозга. Для многих из них характерна фоновая импульсная активность, частота которой возрастает, например, по мере повышения давления в мочевом пузыре.
Представляет собой систему тканей и органов, построенных из нервной ткани. В ней выделяют:
Центральный отдел: головной и спинной мозг
Периферический отдел: автономные и чувствительные ганглии, периферические нервы, нервные окончания.
Существует также подразделение на:
Соматический (анимальный, цереброспинальный) отдел;
Вегетативный (автономный) отдел: симпатическая и парасимпатическая части.
Нервную систему формируют следующие эмбриональные источники: нервная трубка, нервный гребень (ганглиозная пластинка) и эмбриональные плакоды. Тканевые элементы оболочек являются мезенхимными производными. На стадии замыкания нейропоров передний конец трубки значительно расширяется, боковые стенки утолщаются, образуя зачатки трех мозговых пузырей. Лежащий краниально пузырь образует передний мозг, средний пузырь - средний мозг, а из третьего пузыря, который переходит в закладку спинного мозга, развивается задний (ромбовидный) мозг. Вскоре после этого нервная трубка изгибается почти под прямым углом, и посредством борозд-сужений первый пузырь разделяется на конечный и промежуточный отделы, а третий мозговой пузырь - на продолговатый и задний отделы мозга. Производные среднего и заднего мозговых пузырей образуют ствол мозга и являются древними образованиями; в них сохраняется сегментарный принцип строения, который исчезает в производных промежуточного и конечного мозга. В последних концентрируются интегративные функции. Так формируются пять отделов мозга: конечный и промежуточный мозг, средний, продолговатый и задний мозг (у человека это происходит примерно в конце 4-й нед эмбрионального развития). Конечный мозг формирует два полушария большого мозга.
В эмбриональном гисто- и органогенезе нервной системы развитие разных отделов мозга происходит с разной скоростью (гетерохронно). Раньше формируются каудальные отделы центральной нервной системы (спинной мозг, ствол мозга); время окончательного формирования структур головного мозга сильно варьирует. В ряде отделов головного мозга это происходит после рождения (мозжечок, гиппокамп, обонятельная луковица); в каждом отделе мозга существуют пространственно-временные градиенты формирования нейронных популяций, которые образуют уникальную структуру нервного центра.
Спинной мозг представляет собой часть центральной нервной системы, в структуре которой наиболее отчетливо сохраняются черты эмбриональных стадий развития мозга позвоночных: трубчатый характер строения и сегмен-тарность. В боковых отделах нервной трубки быстро возрастает масса клеток, тогда как дорсальная и вентральная ее части не увеличиваются в объеме и сохраняют эпендимный характер. Утолщенные боковые стенки нервной трубки делятся продольной бороздой на дорсальную - крыльную, и вентральную - основную пластинку. На этой стадии развития в боковых стенках нервной трубки можно выделить три зоны: эпендиму, выстилающую центральный канал, промежуточную (плащевой слой) и маргинальную (краевую вуаль). Из плащевого слоя в дальнейшем развивается серое вещество спинного мозга, а из краевой вуали - его белое вещество. Нейробласты передних столбов дифференцируются в мотонейроны (двигательные нейроны) ядер передних рогов. Их аксоны выходят из спинного мозга и образуют передние корешки спинномозговых нервов. В задних столбах и промежуточной зоне развиваются различные ядра вставочных (ассоциативных) клеток. Их аксоны, поступая в белое вещество спинного мозга, входят в состав различных проводящих пучков. В задние рога входят центральные отростки чувствительных нейронов спинномозговых узлов.
Одновременно с развитием спинного мозга закладываются спинномозговые и периферические узлы автономной нервной системы. Исходным материалом для них служат стволовые клеточные элементы нервного гребня, которые путем дивергентной дифференцировки развиваются в нейробла-стическом и глиобластическом направлениях. Часть клеток нервного гребня мигрирует на периферию в места локализации узлов автономной нервной системы, параганглиев, нейроэндокринных клеток APUD-серии и хромаффинной ткани.
Периферическая нервная система.
Периферическая нервная система объединяет периферические нервные узлы, стволы и окончания.
Нервные ганглии (узлы) – структуры, образованные скоплениями нейронов вне ЦНС, - разделяются на чувствительные и автономные (вегетативные). Чувствительные ганглии содержат псевдоуниполярные или биполярные (в спиральном и вестибулярном ганглиях) афферентные нейроны и располагаются преимущественно по ходу задних корешков спинного мозга (чувствительные узлы спинномозговых нервов) и некоторых черепно-мозговых нервов. Чувствительные ганглии спинномозговых нервов имеют веретеновидную форму и покрыты капсулой из плотной волокнистой соединительной ткани. По периферии ганглия находятся плотные скопления тел псевдоуниполярных нейронов, а центральная часть занята их отростками и расположенными между ними тонкими прослойками эндоневрия, несущими сосуды. Автономные нервные ганглии образованы скоплениями мультиполярных нейронов, на которых многочисленные синапсы образуют преганглионарные волокна – отростки нейронов, чьи тела лежат в ЦНС.
Нерв. Строение и регенерация. Спинномозговые ганглии. Морфофункциональная характеристика.
Нервы (нервные стволы) связывают нервные центры головного и спинного мозга с рецепторами и рабочими органами. Они образованы пучками миелиновых и безмиелиновых волокон, которые объединены соединительнотканными компонентамии (оболочками): эндоневрием, периневрием и эпиневрием. Большинство нервов являются смешанными, т.е. включают афферентные и эфферентные волокна.
Эндоневрий – тонкие прослойки рыхлой волокнистой соединительной ткани с мелкими кровеносными сосудами, окружающие отдельные нервные волокна и связывающие их в единый пучок. Периневрий – оболочка, покрывающая каждый пучок нервных волокон снаружи и отдающая перегородки вглубь пучка. Он имеет пластинчатое строение и образов концентрическими пластами уплощённых фиброблатстоподобных клеток, связанных плотными и щелевыми соединениями. Между слоями клеток в пространствах, заполненных жидкостью, располагаются компоненты базальной мембраны и продольно ориентированные коллагеновые волокна. Эпиневрий – наружная оболочка нерва, связывающая воедино пучки нервных волокон. Он состоит из плотной волокнистой соединительной ткани, содержащей жировые клетки, кровеносные и лимфатические сосуды.
Спинной мозг. Морфофункциональная характеристика. Развитие. Строение серого и белого вещества. Нейронный состав.
Спинной мозг состоит из двух симметричных половин, отграниченных друг от друга спереди глубокой серединной щелью, а сзади – соединительнотканной перегородкой. Внутренняя часть органа темнее - это его серое вещество. На периферии спинного мозга располагается более светлое белое вещество. Серое вещество спинного мозга состоит из тел нейронов, безмиелиновых и тонких миелиновых волокон и нейроглии. Основной составной частью серого вещества, отличающей его от белого, являются мультиполярные нейроны. Выступы серого вещества принято называть рогами. Различают передние, или вентральные, задние, или дорсальные, и боковые, или латеральные, рога. В процессе развития спинного мозга из нервной трубки образуются нейроны, группирующиеся в 10 слоях, или в пластинах. Для человека характерна
следующая архитектоникауказанных пластин: I-V пластины соответствуют задним рогам, VI-VII пластины - промежуточной зоне, VIII-IX пластины - передним рогам, X пластина - зона околоцентрального канала. Серое вещество мозга состоит из мультиполярных нейронов трех типов. Первый тип нейронов является филогенетически более древним и характеризуется немногочисленными длинными, прямыми и слабо ветвящимися дендритами (изодендритический тип). Второй тип нейронов имеет большое число сильно ветвящихся дендритов, которые переплетаются, образуя «клубки» (идиодендритический тип). Третий тип нейронов по степени развития дендритов занимает промежуточное положение между первым и вторым типами. Белое вещество спинного мозга представляет собой совокупность продольно ориентированных преимущественно миелиновых волокон. Пучки нервных волокон, осуществляющие связь между различными отделами нервной системы, называются проводящими путями спинного мозга
Головной мозг. Источники развития. Общая морфофункциональная характеристика больших полушарий. Нейронная организация больших полушарий. Цито- и миелоархитектоника коры больших полушарий головного мозга. Возрастные изменения коры.
В головном мозге различают серое и белое вещество, но распределение этих двух составных частей здесь значительно сложнее, чем в спинном мозге. Большая часть серого вещества головного мозга располагается на поверхности большого мозга и в мозжечке, образуя их кору. Меньшая часть образует многочисленные ядра ствола мозга.
Строение . Кора большого мозга представлена слоем серого вещества. Наиболее сильно развита она в передней центральной извилине. Обилие борозд и извилин значительно увеличивает площадь серого вещества головного мозга.. Различные участки ее, отличающиеся друг от друга некоторыми особенностями расположения и строения клеток (цитоархитектоника), расположения волокон (миелоархитектоника) и функциональным значением, называются полями. Они представляют собой места высшего анализа и синтеза нервных импульсов. Резко очерченные
границы между ними отсутствуют. Для коры характерно расположение клеток и волокон слоями. Развитие коры больших полушарий (неокортекса) человека в эмбриогенезе происходит из вентрикулярной герминативной зоны конечного мозга, где расположены малоспециализированные пролиферирующие клетки. Из этих клеток дифференцируются нейроциты неокортекса. При этом клетки утрачивают способность к делению и мигрируют в формирующуюся корковую пластинку. Вначале в корковую пластинку поступают нейроциты будущих I и VI слоев, т.е. наиболее поверхностного и глубокого слоев коры. Затем в нее встраиваются в направлении изнутри и кнаружи последовательно нейроны V, IV, III и II слоев. Этот процесс осуществляется за счет образования клеток в небольших участках вентрикулярной зоны в различные периоды эмбриогенеза (гетерохрон-но). В каждом из этих участков образуются группы нейронов, последовательно выстраивающихся вдоль одного или нескольких волокон
радиальной глии в виде колонки.
Цитоархитектоника коры большого мозга. Мультиполярные нейроны коры весьма разнообразны по форме. Среди них можно выделить пирамидные, звездчатые, веретенообразные, паукообразные и горизонтальные нейроны. Нейроны коры расположены нерезко отграниченными слоями. Каждый слой характеризуется преобладанием какого-либо одного вида клеток. В двигательной зоне коры различают 6 основных слоев: I - молекулярный, II - наружный зернистый, III - nuрамидных нейронов, IV - внутренний зернистый, V - ганглионарный, VI - слой полиморфных клеток. Молекулярный слой коры содержит небольшое количество мелких ассоциативных клеток веретеновидной формы. Их нейриты проходят параллельно поверхности мозга в составе тангенциального сплетения нервных волокон молекулярного слоя. Наружный зернистый слой образован мелкими нейронами, имеющими округлую, угловатую и пирамидальную форму, и звездчатыми нейроцитами. Дендриты этих клеток поднимаются в молекулярный слой. Нейриты или уходят в белое вещество, или, образуя дуги, также поступают в тангенциальное сплетение волокон молекулярного слоя. Самый широкий слой коры большого мозга - пирамидный. От верхушки пирамидной клетки отходит главный дендрит, который располагается в молекулярном слое. Нейрит пирамидной клетки всегда отходит от ее основания. Внутренний зернистый слой образован мелкими звездчатыми нейронами. В его состав входит большое количество горизонтальных волокон. Ганглионарный слой коры образован крупными пирамидами, причем область прецентральной извилины содержит гигантские пирамиды.
Слой полиморфных клеток образован нейронами различной формы.
Миелоархитектоника коры . Среди нервных волокон коры полушарий большого мозга можно выделить ассоциативные волокна, связывающие отдельные участки коры одного полушария, комиссуральные, соединяющие кору различных полушарий, и проекционные волокна, как афферентные, так и эфферентные, которые связывают кору с ядрами низших отделов центральной
нервной системы.
Возрастные изменения . На 1-м году жизни наблюдаются типизация формы пирамидных и звездчатых нейронов, их увеличение, развитие дендритных и аксонных арборизаций, внутриансамблевых связей по вертикали. К 3 годам в ансамблях выявляются «гнездные» группировки нейронов, более четко сформированные вертикальные дендритные пучки и пучки радиарных волокон. К 5-6 годам нарастает полиморфизм нейронов; усложняется система внутриансамблевых связей по горизонтали за счет роста в длину и разветвлений боковых и базальных дендритов пирамидных нейронов и развития боковых терминалей их апикальных дендритов. К 9-10 годам увеличиваются клеточные группировки, значительно усложняется структура короткоаксонных нейронов, и расширяется сеть аксонных коллатералей всех форм интернейронов. К 12-14 годам в ансамблях четко обозначаются специализированные формы пирамидных нейронов, все типы интернейронов достигают высокого уровня дифференцировки. К 18 годам ансамблевая организация коры по основным параметрам своей архитектоники достигает уровня таковой у взрослых.
Мозжечок. Строение и морфофункциональная характеристика. Нейронный состав коры мозжечка, глиоциты. Межнейронные связи.
Мозжечок . Представляет собой центральный орган равновесия и координации движений. Он связан со стволом мозга афферентными и эфферентными проводящими пучками, образующими в совокупности три пары ножек мозжечка. На поверхности мозжечка много извилин и бороздок, которые значительно увеличивают ее площадь. Борозды и извилины создают на разрезе
характерную для мозжечка картину «древа жизни». Основная масса серого вещества в мозжечке располагается на поверхности и образует его кору. Меньшая часть серого вещества лежит глубоко в белом веществе в виде центральных ядер. В центре каждой извилины имеется тонкая прослойка
белого вещества, покрытая слоем серого вещества - корой. В коре мозжечка различают три слоя: наружный - молекулярный, средний - ганглионарный слой, или слой грушевидных нейронов, и внутренний -зернистый. Ганглиозный слой содержит грушевидные нейроны. Они имеют нейриты, которые, покидая кору мозжечка, образуют начальное звено его эфферентных
тормозных путей. От грушевидного тела в молекулярный слой отходят 2-3 дендрита, которые пронизывают всю толщу молекулярного слоя. От основания тел этих клеток отходят нейриты, проходящие через зернистый слой коры мозжечка в белое вещество и заканчивающиеся на клетках ядер мозжечка. Молекулярный слой содержит два основных вида нейронов: кор-зинчатые и звездчатые. Корзинчатые нейроны находятся в нижней трети молекулярного слоя. Их тонкие длинные дендриты ветвятся преимущественно в плоскости, расположенной поперечно к извилине. Длинные нейриты клеток всегда идут поперек извилины и параллельно поверхности над грушевидными нейронами. Звездчатые нейроны лежат выше корзинчатых и эывают двух типов. Мелкие звездчатые нейроны снабжены тонкими короткими дендритами и слаборазветвленными нейритами, образующими синапсы. Крупные звездчатые нейроны имеют длинные и сильно разветвленные дендриты и нейриты. Зернистый слой. Первым типом клеток этого слоя можно считать зерновидные нейроны, или клетки-зерна. Клетка имеет 3-4 коротких дендрита,
заканчивающихся в этом же слое концевыми ветвлениями в виде лапки птицы. Нейриты клеток-зерен проходят в молекулярный слой и в нем делятся на две ветви, ориентированные параллельно поверхности коры вдоль извилин мозжечка. Вторым типом клеток зернистого слоя мозжечка являются тормозные большие звездчатые нейроны. Различают два вида таких клеток: с короткими и длинными нейритами. Нейроны с короткими нейритами лежат вблизи ганглионарного слоя. Их разветвленные дендриты распространяются в молекулярном слое и образуют синапсы с параллельными волокнами - аксонами клеток-зерен. Нейриты направляются в зернистый слой к клубочкам мозжечка и заканчиваются синапсами на концевых ветвлениях дендритов клеток-зерен.
Немногочисленные звездчатые нейроны с длинными нейритами имеют обильно ветвящиеся в зернистом слое дендриты и нейриты, выходящие в белое вещество. Третий тип клеток составляют веретеновидные горизонтальные клетки. Они имеют небольшое вытянутое тело, от которого в обе стороны отходят длинные горизонтальные дендриты, заканчивающиеся в ганглионарном и зернистом слоях. Нейриты же этих клеток дают коллатерали в зернистый слой и уходят в
белое вещество. Глиоциты . Кора мозжечка содержит различные глиальные элементы. В зернистом слое имеются волокнистые и протоплазматические астроциты. Ножки отростков волокнистых астроцитов образуют периваскулярные мембраны. Во всех слоях в мозжечке имеются олигодендроциты. Особенно богаты этими клетками зернистый слой и белое вещество мозжечка. В ганглионарном слое между грушевидными нейронами лежат глиальные клетки с темными ядрами. Отростки этих клеток направляются к поверхности коры и образуют глиальные волокна молекулярного слоя мозжечка. Межнейрональные связи . Афферентные волокна, поступающие в кору мозжечка, представлены двумя видами - моховидными и так называемыми лазящими волокнами. Моховидные волокна идут в составе оливомозжечкового и мостомозжечкового путей и опосредованно через клетки-зерна оказывают на грушевидные клетки возбуждающее действие.
Лазящие волокна поступают в кору мозжечка, по-видимому, по спинно-мозжечковому и вестибуломозжечковому путям. Они пересекают зернистый слой, прилегают к грушевидным нейронам и стелются по их дендритам, заканчиваясь на их поверхности синапсами. Лазящие волокна передают возбуждение непосредственно грушевидным нейронам.
Автономная (вегетативная) нервная система. Общая морфофункциональная характеристика. Отделы. Строение экстрамуральных и интрамуральных ганглиев.
ВНС делится на симпатическую и парасимпатическую. Обе системы одновременно принимают участие в иннервации органов и оказывают на них противоположное влияние. Состоит из центральных отделов, представленных ядрами серого вещества головного и спинного мозга, и периферических: нервных стволов, узлов (ганглиев) и сплетений.
Интрамуральные ганглии и связанные с ними проводящие пути ввиду их высокой автономии, сложности организации и особенностей медиаторного обмена выделяют в самостоятельный метасимпатический отдел автономной НС. Выделяют нейроны трёх типов:
Длинноаксонные эфферентные нейроны (клетки I типа Догеля) с короткими дендритами и длинным аксоном, идущим за пределы узла к клеткам рабочего органа, на которых он образует двигательные или секреторные окончания.
Равноотросчатые афферентные нейроны (клетки II типа Догеля) содержат длинные дендриты и аксон, уходящий за пределы данного ганглия в соседние и образующий синапсы на клетках I и III типов. Входят в качестве рецепторного звена в состав местных рефлекторных дуг, которые замыкаются без захода нервного импульса в ЦНС.
Ассоциативные клетки (клетки III типа Догеля) – местные вставочные нейроны, соединяющие своими отростками несколько клеток I и II типов. Дендриты этих клеток не выходят за пределы узла, а аксоны направляются в другие узлы, образуя синапсы на клетках I типа.