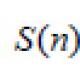Изменение энтропии в биологических системах. Энтропия биологических систем Гибель живых организмов сопровождается энтропии
Отправить свою хорошую работу в базу знаний просто. Используйте форму, расположенную ниже
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Размещено на http://allbest.ru
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РФ
Федеральное государственное автономное образовательное учреждение высшего профессионального образования
«САНКТ-ПЕТЕРБУРГСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ АЭРОКОСМИЧЕСКОГО ПРИБОРОСТРОЕНИЯ»
КАФЕДРА ЭКОНОМИЧЕСКОЙ БЕЗОПАСНОСТИ
Энтропия в живых системах
по дисциплине: Концепции современного естествознания
Выполнила: студент
Руководитель:
д-р экон. наук,
профессор А. В. Самойлов
Санкт-Петербург 2014
Электронная схема жизни
Энтропия жизни
Диагностические технологии, основанные на достижениях квантовой биофизики
Заключение
Список литературы
Ведение
Методы исследования функционального состояния человека путем регистрации электрооптических параметров кожного покрова можно разделить на две условные группы по характеру вовлекаемых биофизических процессов. К первой группе относятся "медленные" методы, время измерения в которых составляет более 1 с.
При этом под влиянием приложенных потенциалов в тканях стимулируются ионно-деполяризационные токи и основной вклад в измеряемый сигнал вносит ионная компонента . "Быстрые" методы, время измерения в которых составляет менее 100 мс, основаны на регистрации физических процессов, стимулированных электронной компонентой тканевой проводимости.
Подобные процессы описываются в основном квантово-механическими моделями, поэтому их можно обозначить как методы квантовой биофизики.
К последним относятся методы регистрации стимулированной и собственной люминесценции, а также метод стимулированной электронной эмиссии с усилением в газовом разряде (метод газоразрядной визуализации). Рассмотрим более детально биофизические и энтропийные механизмы реализации методов квантовой биофизики.
Электронная схема жизни
"Я глубоко убежден, что мы никогда не сможем понять сущность жизни, если ограничимся молекулярным уровнем… Удивительная тонкость биологических реакций обусловлена подвижностью электронов и объяснима только с позиций квантовой механики". А. Сент-Дьердьи, 1971
Электронная схема жизни - круговорот и превращение энергии в биологических системах. Фотоны солнечного света поглощаются молекулами хлорофиллов, сосредоточенных в мембранах хлоропластов органоидов зеленых растений.
Поглощая свет, электроны хлорофиллов приобретают дополнительную энергию и переходят из основного в возбужденное состояние. Благодаря упорядоченной организации белково-хлорофиллового комплекса, который носит название фотосистемы (ФС), возбужденный электрон не тратит энергию на тепловые превращения молекул, а приобретает способность преодолевать электростатическое отталкивание, хотя расположенное рядом с ним вещество имеет более высокий электронный потенциал, чем хлорофилл. В результате возбужденный электрон переходит на это вещество.
После потери своего электрона хлорофилл имеет свободную электронную вакансию. И он отбирает электрон у окружающих молекул, причем донором могут служить вещества, электроны которых имеют меньшую энергию, чем электроны хлорофилла. Этим веществом является вода.
Отбирая электроны у воды, фотосистема окисляет ее до молекулярного кислорода. Так атмосфера Земли непрерывно обогащается кислородом.
При переносе подвижного электрона по цепи структурно связанных между собой макромолекул он тратит свою энергию на анаболические и катаболические процессы в растениях, а при соответствующих условиях, и у животных. По современным представлениям межмолекулярный перенос возбужденного электрона происходит по механизму туннельного эффекта в сильном электрическом поле.
Хлорофиллы служат промежуточной ступенькой в потенциальной яме между донором и акцептором электронов. Они принимают электроны от донора с низким энергетическим уровнем и за счет энергии солнца возбуждают их настолько, что они могут переходить на вещество с более высоким электронным потенциалом, чем донор.
Это единственная, хотя и многоступенчатая световая реакция в процессе фотосинтеза. Дальнейшие аутотрофные биосинтетические реакции не нуждаются в свете. Они происходят в зеленых растениях за счет энергии, заключенной в электронах, принадлежащих НАДФН и АТФ. За счет колоссального притока электронов из двуокиси углерода, воды, нитратов, сульфатов и прочих сравнительно простых веществ создаются высокомолекулярные соединения: углеводы, белки, жиры, нуклеиновые кислоты.
Эти вещества служат основными питательными веществами для гетеротрофов. В ходе катаболических процессов, также обеспечиваемых электрон-транспортными системами, освобождаются электроны примерно в таком же количестве, в каком они захватывались органическими веществами при их фотосинтезе.
Электроны, освобождаемые при катаболизме, переносятся на молекулярный кислород дыхательной цепью митохондрий. Здесь окисление сопряжено с фосфорилированием - синтезом АТФ посредством присоединения к АДФ остатка фосфорной кислоты (то есть фосфорилирования АДФ). Этим обеспечивается энергоснабжение всех процессов жизнедеятельности животных и человека.
Находясь в клетке, биомолекулы "живут", обмениваясь энергией и зарядами, а значит, информацией, благодаря развитой системе делокализованных р-электронов. Делокализация означает, что единое облако р-электронов распределено определенным образом по всей структуре молекулярного комплекса. Это позволяет им мигрировать не только в пределах своей молекулы, но и переходить с молекулы на молекулу, если они структурно объединены в ансамбли. Явление межмолекулярного переноса было открыто J. Weiss в 1942 г., а квантовомеханическую модель этого процесса разработал в 1952-1964 гг R.S. Mulliken.
Вместе с тем важнейшая миссия р-электронов в биологических процессах связана не только с их делокализацией, но и с особенностями энергетического статуса: разность энергий основного и возбужденного состояний для них значительно меньше, чем у р-электронов и примерно равна энергии фотона hн.
Благодаря этому именно р-электроны способны аккумулировать и конвертировать солнечную энергию, за счет чего с ними связано все энергообеспечение биологических систем. Поэтому р-электроны принято называть "электронами жизни".
Сопоставляя шкалы восстановительных потенциалов компонентов систем фотосинтеза и дыхательной цепи, нетрудно убедиться в том, что солнечная энергия, конвертированная р-электронами при фотосинтезе, затрачивается преимущественно на клеточное дыхание (синтез АТФ). Так, за счет поглощения двух фотонов в хлоропласте р-электроны переносятся от Р680 до ферредоксина, увеличивая свою энергию примерно на 241 кДж/моль. Ее небольшая часть расходуется при переносе р-электронов с ферредоксина на НАДФ. В результате синтезируются вещества, которые затем становятся пищей для гетеротрофов и превращаются в субстраты клеточного дыхания. В начале дыхательной цепи запас свободной энергии р-электронов составляет 220 кДж/моль. Значит, до этого энергия р-электронов понизилась всего на 20 кДж/моль. Следовательно, более 90% солнечной энергии, запасенной р-электронами в зеленых растениях, доносится ими до дыхательной цепи митохондрий животных и человека.
Конечным продуктом окислительно-восстановительных реакций в дыхательной цепи митохондрий является вода. Она обладает наименьшей свободной энергией из всех биологически важных молекул. Говорят, будто с водой организм выделяет электроны, лишенные энергии в процессах жизнедеятельности. На самом деле запас энергии в воде отнюдь не нулевой, но вся энергия заключена в у-связях и не может быть использована для химических превращений в организме при температуре тела и других физико-химических параметрах организма животных и человека. В этом смысле химическую активность воды принимают за точку отсчета (нулевой уровень) на шкале химической активности.
Из всех биологически важных веществ вода обладает самым высоким ионизационным потенциалом - 12,56 эВ. У всех молекул биосферы ионизационные потенциалы ниже этой величины, диапазон величин находится примерно в пределах 1 эВ (от 11,3 до 12,56 эВ).
Если принять ионизационный потенциал воды за точку отсчета реакционной способности биосферы, то можно построить шкалу биопотенциалов. Биопотенциал каждого органического вещества имеет вполне определенное значение - он соответствует энергии, которая освобождается при окислении данного соединения до воды.
Размерность БП - это размерность свободной энергии соответствующих веществ (в ккал). И хотя 1 эВ = 1,6 10 -19 Дж, при переходе от шкалы ионизационных потенциалов к шкале биопотенциалов нужно учитывать число Фарадея и разность стандартных восстановительных потенциалов между редокс-парой данного вещества и редокс-парой О 2 /H 2 O.
Благодаря поглощению фотонов электроны достигают наивысшего биопотенциала в фотосистемах растений. С этого высокого энергетического уровня они дискретно (по ступенькам) спускаются на самый низкий в биосфере энергетический уровень - уровень воды. Энергия, отдаваемая электронами на каждой ступеньке этой лестницы, превращается в энергию химических связей и таким образом движет жизнью животных и растений. Электроны воды связываются растениями, а клеточное дыхание вновь порождает воду. Этот процесс образует электронный кругооборот в биосфере, источником которого служит солнце.
Еще одним классом процессов, являющихся источником и резервуаром свободной энергии в организме, являются окислительные процессы, протекающие в организме с участием активных форм кислорода (АФК). АФК - это высоко реакционно-способные химические частицы, к которым относят содержащие кислород свободные радикалы (О 2 3 / 4 ·, HО 2 ·, НО·, NO·, ROO), а также молекулы, способные легко продуцировать свободные радикалы (синглетный кислород, O 3 , ONOOH, HOCl, H 2 O 2 , ROOH, ROOR). В большинстве посвященных АФК публикаций, обсуждаются вопросы, связанные с их патогенным действием, поскольку долгое время считалось, что АФК появляются в организме при нарушениях нормального метаболизма, а в ходе инициированных свободными радикалами цепных реакций неспецифически повреждаются молекулярные компоненты клетки.
Однако сейчас стало ясно, что генерирующие супероксид ферменты имеются практически у всех клеток и что многие нормальные физиологические реакции клеток коррелируют с увеличением продукции АФК . АФК генерируются и в ходе постоянно протекающих в организме неферментативных реакций. По минимальным оценкам, в состоянии покоя при дыхании человека и животных на производство АФК идет до 10-15% кислорода, а при повышении активности эта доля существенно возрастает. При этом стационарный уровень АФК в органах и тканях в норме очень низок благодаря повсеместной распространенности мощных ферментативных и неферментативных устраняющих их систем. Вопрос о том, зачем организм столь интенсивно продуцирует АФК с тем, чтобы немедленно от них избавиться, в литературе пока не обсуждается.
Установлено, что адекватные реакции клетки на гормоны, нейромедиаторы, цитокины, на физические факторы (свет, температура, механические воздействия) требуют определенного содержании АФК в среде. АФК и сами могут вызывать в клетках те же реакции, что развиваются под действием биорегуляторных молекул - от активации или обратимого ингибирования ферментативных систем до регуляции активности генома. Биологическая активность так называемых аэроионов, оказывающих выраженное терапевтическое действие на широкий круг инфекционных и неинфекционных заболеваний , обусловлена тем, что они представляют собой свободные радикалы (О 2 3/4 ·) . Расширяется применение в терапевтических целях и других АФК - озона и перекиси водорода.
Важные результаты были получены в последние годы профессором Московского государственного университета В.Л. Воейковым . На основании большого объема экспериментальных данных по исследованию сверхслабого свечения цельной неразведенной крови человека было установлено, что в крови непрерывно протекают реакции с участием АФК, в ходе которых генерируются электронно-возбужденные состояния (ЭВС).
Аналогичные процессы могут быть инициированы и модельных водных системах, содержащих аминокислоты и компоненты, способствующие медленному окислению аминокислот, в условиях близких к физиологическим. Энергия электронного возбуждения может излучательно и безызлучательно мигрировать в водных модельных системах и в крови, и использоваться как энергия активации для интенсификации процессов, порождающих ЭВС, в частности, за счет индукции вырожденного разветвления цепей.
Процессы с участием АФК, протекающие в крови и в водных системах, проявляют признаки самоорганизации, выражающиеся в их колебательном характере, устойчивости к действию интенсивных внешних факторов при сохранении высокой чувствительности к действию факторов низкой и сверхнизкой интенсивности. Это положение закладывает основу для объяснения многих эффектов, используемых в современной низкоинтенсивной терапии.
Полученные В.Л. Воейковым результаты демонстрируют еще один механизм генерации и утилизации ЭВС в организме, на этот раз в жидких средах. Развитие представлений, изложенных в данной главе, позволит обосновать биофизические механизмы генерации и транспорта энергии в биологических системах.
Энтропия жизни
В термодинамическом отношении открытые (биологические) системы в процессе функционирования проходят через ряд неравновесных состояний, что, в свою очередь, сопровождается изменением термодинамических переменных.
Поддержание неравновесных состояний в открытых системах возможно лишь за счет создания в них потоков вещества и энергии, что говорит о необходимости рассмотрения параметров таких систем как функции времени.
Изменение энтропии открытой системы может происходить за счет обмена с внешней средой (d e S) и за счет роста энтропии в самой системе вследствие внутренних необратимых процессов (d i S > 0). Э. Шредингер ввел понятие, что общее изменение энтропии открытой системы складывается из двух частей:
dS = d e S + d i S.
Продифференцировав это выражение, получим:
dS/dt = d e S/dt + d i S/dt.
Полученное выражение означает, что скорость изменения энтропии системы dS/dt равна скорости обмена энтропией между системой и окружающей средой плюс скорость возникновения энтропии внутри системы.
Член d e S/dt , учитывающий процессы обмена энергией с окружающей средой, может быть и положительным, и отрицательным, так что при d i S > 0 общая энтропия системы может как возрастать, так и убывать.
Отрицательная величина d e S/dt < 0 соответствует тому, что отток положительной энтропии от системы во внешнюю среду превышает приток положительной энтропии извне, так что в результате общая величина баланса обмена энтропией между системой и средой является отрицательной. Очевидно, что скорость изменения общей энтропии системы может быть отрицательной при условии:
dS/dt < 0 if d e S/dt < 0 and |d e S/dt| > d i S/dt.
Таким образом, энтропия открытой системы уменьшается за счет того, что в других участках внешней среды идут сопряженные процессы с образованием положительной энтропии.
Для земных организмов общий энергообмен можно упрощенно представить как образование в фотосинтезе сложных молекул углеводов из СО 2 и Н 2 О с последующей деградацией продуктов фотосинтеза в процессах дыхания. Именно этот энергообмен обеспечивает существование и развитие как отдельных организмов - звеньев в круговороте энергии. Так и жизни на Земле в целом.
С этой точки зрения уменьшение энтропии живых систем в процессе их жизнедеятельности обусловлено в конечном итоге поглощением квантов света фотосинтезирующими организмами, что, однако, с избытком компенсируется образованием положительной энтропии в недрах Солнца. Этот принцип относится и к отдельным организмам, для которых поступление извне питательных веществ, несущих приток "отрицательной" энтропии, всегда сопряжено с продуцированием положительной энтропии при их образовании в других участках внешней среды, так что суммарное изменение энтропии в системе организм + внешняя среда всегда положительно.
При неизменных внешних условиях в частично равновесной открытой системе в стационарном состоянии, близком к термодинамическому равновесию, скорость прироста энтропии за счет внутренних необратимых процессов достигает отличного от нуля постоянного минимального положительного значения.
d i S/dt => A min > 0
Этот принцип минимума прироста энтропии, или теорема Пригожина, представляет собой количественный критерий для определения общего направления самопроизвольных изменений в открытой системе вблизи равновесия.
Это условие можно представить по-другому:
d/dt (d i S/dt) < 0
Это неравенство свидетельствует об устойчивости стационарного состояния. Действительно, если система находится в стационарном состоянии, то она не может самопроизвольно выйти из него за счет внутренних необратимых изменений. При отклонении от стационарного состояния в системе должны произойти внутренние процессы, возвращающие ее к стационарному состоянию, что соответствует принципу Ле-Шателье - устойчивости равновесных состояний. Иными словами, любое отклонение от устойчивого состояния вызовет увеличение скорости продуцирования энтропии.
В целом уменьшение энтропии живых систем происходит за счет свободной энергии, освобождаемой при распаде поглощаемых извне питательных веществ или за счет энергии солнца. Одновременно это приводит к увеличению их свободной энергии.
Таким образом, поток отрицательной энтропии необходим для компенсации внутренних деструктивных процессов и убыли свободной энергии за счет самопроизвольных реакций метаболизма. В сущности, речь идет о круговороте и превращении свободной энергии, за счет которой поддерживается функционирование живых систем.
Диагностические технологии, основанные на достижениях квантовой биофизики
На основании рассмотренных выше представлений был развит целый ряд подходов, позволяющих исследовать прижизненную активность биологических систем.
Это прежде всего спектральные методы, среди которых необходимо отметить методику одновременного измерения собственной флуоресценции НАДН и окисленных флавопротеидов (ФП), развитую коллективом авторов под руководством В.О. Самойлова.
Эта методика основана на использовании оригинальной оптической схемы, разработанной Е.М. Брумбергом, позволяющей одномоментно измерять флуоресценцию НАДН на длине волны л = 460 нм (синий свет) и флуоресценцию ФП на длине волны л = 520-530 нм (желто-зеленый свет) при возбуждении ультрафиолетом (л = 365 нм).
В этой донорно-акцепторной паре донор р-электронов флуоресцирует в восстановленной форме (НАДН), а акцептор - в окисленной (ФП). Естественно, что в покое преобладают восстановленные формы, а при усилении окислительных процессов - окисленные.
Методика была доведена до практического уровня удобных эндоскопических приборов, что позволило разработать раннюю диагностику злокачественных заболеваний желудочно-кишечного тракта, лимфатических узлов в процессе хирургических операций, кожи. Принципиально важной оказалась оценка степени жизнеспособности тканей в процессе хирургических операций для проведения экономной резекции.
Прижизненная флуометрия дает, кроме статических показателей, динамические характеристики биологических систем, так как позволяет проводить функциональные пробы и исследовать зависимость типа "доза-эффект". Это обеспечивает в клинике надежную функциональную диагностику и служит инструментом экспериментального изучения интимных механизмов патогенеза заболеваний.
К направлению квантовой биофизики можно отнести и метод газоразрядной визуализации (ГРВ). Стимулирование эмиссии электронов и фотонов с поверхности кожного покрова происходит за счет коротких (10 мкс) импульсов электромагнитного поля (ЭМП). Как показали измерения при помощи импульсного осциллографа с памятью, во время действия импульса ЭМП развивается серия импульсов тока (и свечения) длительностью примерно 10 нс каждый.
Развитие импульса обусловлено ионизацией молекул газовой среды за счет эмитированных электронов и фотонов, срыв импульса связан с процессами зарядки диэлектрической поверхности и возникновением градиента ЭМП, направленного противоположно исходному полю. При подаче серии стимулирующих импульсов ЭМП с частотой следования 1000 Гц эмиссионные процессы развиваются в течение времени действия каждого импульса.
Телевизионное наблюдение временной динамики свечения участка кожного покрова диаметром несколько миллиметров и покадровое сравнение картин свечения в каждом импульсе напряжения свидетельствует о возникновении эмиссионных центров практически в одних и тех же точках кожи. биологический энтропия белковый
За столь короткое время - 10 нс - ионно-деполизационные процессы в ткани развиться не успевают, поэтому ток может быть обусловлен транспортом электронов по структурным комплексам кожи или иной исследуемой биологической ткани, включенной в цепь протекания импульсного электрического тока. Биологические ткани принято разделять на проводники (в первую очередь биологические проводящие жидкости) и диэлектрики.
Для объяснения эффектов стимулированной электронной эмиссии необходимо рассматривать механизмы транспорта электронов по непроводящим структурам. Неоднократно высказывались идеи применить к биологическим тканям модель полупроводниковой проводимости. Полупроводниковая модель миграции электрона на большие межмолекулярные расстояния по зоне проводимости в кристаллической решетке хорошо известна и активно применяется в физике и технике.
В соответствии с современными представлениями, полупроводниковая концепция не получила подтверждения для биологических систем. В настоящее время наибольшее внимание в этой области привлекает к себе концепция туннельного транспорта электронов между отдельными белковыми молекулами-переносчиками, отделенными друг от друга энергетическими барьерами.
Процессы туннельного транспорта электронов хорошо экспериментально изучены и промоделированы на примере переноса электронов по белковой цепи. Туннельный механизм обеспечивает элементарный акт переноса электрона между донорно-акцепторными группами в белке, находящимися друг от друга на расстоянии порядка 0,5 - 1,0 нм. Однако существует много примеров, когда электрон переносится в белке на гораздо большие расстояния.
Существенно, что при этом перенос происходит не только в пределах одной молекулы белка, но может включать взаимодействие разных белковых молекул. Так, в реакции переноса электрона между цитохромами с и цитохромом-оксидазой и цитохромом b5 оказалось, что расстояние между геммами взаимодействующих белков составляет более 2,5 нм . Характерное время переноса электрона составляет 10 -11 - 10 -6 с, что соответствует времени развития единичного эмиссионного акта в методе ГРВ.
Проводимость белков может носить примесный характер. По данным экспериментов, значение подвижности u [м 2 /(В см)] в переменном электрическом поле составили для цитохрома ~ 1*10 -4 , для гемоглобина ~ 2*10 -4 . В целом оказалось, что для большинства белков проводимость осуществляется в результате прыжков электронов между локализованными донорными и акцепторными состояниями, разделенными расстояниями в десятки нанометров. Лимитирующей стадией в процессе переноса является не движение заряда по токовым состояниям, а релаксационные процессы в доноре и акцепторе.
В последние годы удалось рассчитать реальные конфигурации такого рода "электронных троп" в конкретных белках. В этих моделях белковая среда между донором и акцептором разбивается на отдельные блоки, связанные между собой ковалентными и водородными связями, а также невалентными взаимодействиями на расстоянии порядка Ван-дер-ваальсовых радиусов. Электронная тропа, таким образом, представляется комбинацией тех атомных электронных орбиталей, которые дают наибольший вклад в величину матричного элемента взаимодействия волновых функций компонентов.
В то же время общепризнанно, что конкретные пути переноса электрона не носят строго фиксированный характер. Они зависят от конфирмационного состояния белковой глобулы и могут соответственно меняться в различных условиях.
В работах Маркуса был развит подход, в котором рассматривается не одна-единственная оптимальная траектория переноса в белке, а их набор. При вычислении константы переноса принимались во внимание орбитали целого ряда электронно-взаимодействующих атомов аминокислотных остатков белка между донорной и акцепторной группами, которые дают наибольший вклад в суперобменное взаимодействие. Оказалось, что для отдельных белков получаются более точные линейные зависимости, чем при учете одной-единственной траектории.
Трансформация электронной энергии в биоструктурах связана не только с переносом электронов, но и с миграцией энергии электронного возбуждения, которая не сопровождается отрывом электрона от молекулы донора. Наиболее важными для биологических систем, по современным представлениям, оказываются индуктивно-резонансный, обменно-резонансный и экситонный механизмы переноса электронного возбуждения. Эти процессы оказываются важными при рассмотрении процессов переноса энергии по молекулярным комплексам, как правило, не сопровождающихся переносом заряда.
Заключение
Рассмотренные представления показывают, что основным резервуаром свободной энергии в биологических системах являются электронно-возбужденные состояния сложных молекулярных комплексов. Эти состояния непрерывно поддерживаются за счет кругооборота электронов в биосфере, источником которого является солнечная энергия, а основным "рабочим веществом" - вода. Часть состояний тратится на обеспечение текущего энергоресурса организма, часть может запасаться впредь, подобно тому, как это происходит в лазерах после поглощения импульса накачки.
Протекание импульсного электрического тока в непроводящих биологических тканях может обеспечиваться за счет межмолекулярного переноса возбужденных электронов по механизму туннельного эффекта с активированным перескоком электронов в контактной области между макромолекулами.
Таким образом, можно предположить, что формирование специфических структурно-белковых комплексов в толще эпидермиса и дермиса кожи обеспечивает формирование каналов повышенной электронной проводимости, экспериментально измеряемых на поверхности эпидермиса как электропунктурные точки.
Гипотетически можно предположить наличие таких каналов и в толще соединительной ткани, что может быть ассоциировано с "энергетическими" меридианами. Иными словами, понятие переноса "энергии", характерное для представлений Восточной медицины и режущее слух человеку с европейским образованием, может быть ассоциировано с транспортом электронно-возбужденных состояний по молекулярным белковым комплексам.
При необходимости совершения физической или умственной работы в данной системе организма электроны, распределенные в белковых структурах, транспортируются в данное место и обеспечивают процесс окислительного фосфорилирования, то есть энергетического обеспечения функционирования локальной системы.
Таким образом, организм формирует электронное "энергетическое депо", поддерживающее текущее функционирование и являющееся базисом для совершения работы, требующей мгновенной реализации огромных энергоресурсов или протекающей в условиях сверхбольших нагрузок, характерных, например, для профессионального спорта.
Стимулированная импульсная эмиссия также развивается в основном за счет транспорта делокализованных р-электронов, реализуемых в электрически непроводящей ткани путем туннельного механизма переноса электронов. Это позволяет предположить, что метод ГРВ позволяет косвенным образом судить об уровне энергетических запасов молекулярного уровня функционирования структурно-белковых комплексов.
Список литературы
1. Goldstein N.I., Goldstein R.H., Merzlyak M.N. 1992. Negative air ions as a source of superoxide. Int. J. Biometeorol., V. 36., pp. 118-122.
2. Khan, A.U. and Wilson T. Reactive Oxygen Species as Second Messengers. Chem. Biol. 1995. 2: 437-445.
3. Koldunov V.V., Kononov D.S., Voeikov V.L. Sustained chemiluminescence oscillations during Maillard reaction proceeding in aqueous solutions of amino acids and monosaccarides. In: Chemilumunescence at the Turn of the Millenium. Stephen Albrecht, Tomas Zimmerman and Herbert Brandl (eds.) SCHWEDA-WEBERDRUCK GmbH, Druckerei & Verlag, Dresden, 2001, pp. 59-64.
4. Mullarkey CJ, Edelstein D, Brownlee M Free radical generation by early glycation products: a mechanism for accelerated atherogenesis in diabetes. Biochem Biophys Res Commun 1990 Dec 31 173:3 932-9
5. Novikov C.N., Voeikov V.L., Asfaramov R.R., Vilenskaya N.D. Comparative study of peculiarities of chemiluminescene in non-diluted human blood and isolated neutrophiles. In: Chemilumunescence at the Turn of the Millenium. Stephen Albrecht, Tomas Zimmerman and Herbert Brandl (eds.) SCHWEDA-WEBERDRUCK GmbH, Druckerei & Verlag, Dresden, 2001, pp. 130-135.
6. Sauer H., Wartenberg M, Hescheler J. (2001) Reactive Oxygen Species as Intracellular Messengers During Cell Growth and Differentiation. Cell Physiol Biochem;11:173-186.
7. Tiller W. On the evolution of Electrodermal Diagnostic Instruments. J of Advancement in Medicine. 1,1, (1988), pp. 41-72.
8. Vlessis AA; Bartos D; Muller P; Trunkey DD Role of reactive O2 in phagocyte-induced hypermetabolism and pulmonary injury. J Appl Physiol, 1995 Jan, 78:1, 112
9. Voeikov V. Reactive Oxygen Species, Water, Photons, and Life. // Rivista di Biologia/Biology Forum 94 (2001), pp. 193-214
10. Воейков В.Л. Благотворная роль активных форм кислорода. // "Российский журнал гастроэнтерологии, гепатологии, колопроктологии" 2001 год, том XI, № 4, С. 128-135.
11. Воейков В.Л. Регуляторные функции активных форм кислорода в крови и в водных модельных системах. Автореферат Диссертация на соискание ученой степени доктора биологических наук. М. МГУ. 2003
12. Коротков К. Г. Основы ГРВ биоэлектрографии. Ст. Петербург. СПбГИТМО. 2001.
13. Лукьянова Л.Д.. Балмуханов Б.С., Уголев А.Т. Кислород-зависимые процессы в клетке и ее функциональное состояние. М.: Наука, 1982
14. Рубин А.Б. Биофизика. М. Книжный дом "Университет". 1999.
15. Самойлов В.О. Электронная схема жизни. Ст. Петербург, Институт физиологии РАН. 2001.
16. Самойлов В.О. Медицинская биофизика. Ленинград. ВМА. 1986.
18. Чижевский А.Л. Аэроионы и жизнь. М. Мысль. 1999
Размещено на Allbest.ru
...Подобные документы
Физический смысл возрастания энтропии. Характеристика самоорганизации в диссипативных структурах. Особенности эволюции в социальных и гуманитарных системах. Сущность процессов взаимопревращения различных видов энергии. Термодинамическое равновесие.
контрольная работа , добавлен 19.04.2015
Понятие и свойства обратной связи, ее распространенность и значение в живой природе, технике и обществе. Сущность теории двойственной связи как важной стороны управления в живых системах. Отличительные признаки положительных и отрицательных связей.
реферат , добавлен 27.06.2010
Современные представления о структуре организации электрон-транспортной цепи митохондрий и молекулярные преобразователи в клетках. Роль нарушений энергетики в развитии патологии. Основные молекулярные механизмы потребления энергии живой клеткой.
контрольная работа , добавлен 23.02.2014
Энтропия как мера неопределенности, мера хаоса, ее физический смысл. Энтропия в термодинамике – мера необратимого рассеивания энергии, является функцией состояния термодинамической системы. Энтропия Вселенной, энтропия и информация, понятие негэнтропии.
реферат , добавлен 24.03.2010
Теплота и энтропия. Сложность понимания физического смысла энтропии. Энтропия Вселенной, теория тепловой смерти. Сфера применения законов термодинамики. Энтропия как функция состояния термодинамической системы для описания эволюции реальных систем.
реферат , добавлен 18.11.2009
Сущность понятия "биоэнергетика". Существенные признаки живого. Внешний и промежуточный обмен веществ и энергии. Метаболизм: понятие, функции. Три стадии катаболических превращений основных питательных веществ в клетке. Отличия катаболизма от анаболизма.
презентация , добавлен 05.01.2014
Проектирование и создание новых биологических систем, не встречающихся в природе. Методы синтеза искусственных органических молекул, играющих определённую роль в живых системах. Генетическая модификация бактерий с помощью технологии рекомбинантных ДНК.
презентация , добавлен 14.11.2016
Общая характеристика дыхательной цепи как системы структурно и функционально связанных трансмембранных белков и переносчиков электронов. Организация дыхательной цепи в митохондриях. Роль дыхательной цепи в улавливании энергии. Задачи и цели ингибиторов.
реферат , добавлен 29.06.2014
Понятие энтропии как меры хаоса, ее принципы и место в истории развития классической физики. Общая характеристика образования структур нарастающей сложности. Анализ взаимосвязи экологии и естествознания. Оценка экологической обстановки в г. Новосибирске.
реферат , добавлен 21.10.2010
Этапы зарождения жизни на Земле, появления первых прокариотических и эукаротических организмов. Процесс эволюции эукариотов от одноклеточного до многоклеточного строения тела. Основные свойства и проявления жизни, закономерности изменения энтропии.
Общепринятая в физике формулировка второго начала термодинамики гласит, что в закрытых системах энергия стремится распределиться равномерно, т.е. система стремится к состоянию максимальной энтропии.
Отличительной же особенностью живых тел, экосистем и биосферы в целом является способность создавать и поддерживать высокую степень внутренней упорядоченности, т.е. состояния с низкой энтропией. Понятие энтропии характеризует ту часть полной энергии системы, которая не может быть использована для производства работы. В отличие от свободной энергии она представляет собой деградированную, отработанную энергию. Если обозначить свободную энергию через F и энтропию через S , то полная энергия системы Е будет равна:
E = F + ST ;
где Т — абсолютная температура по Кельвину.
По определению физика Э. Шредингера: «жизнь — упорядоченное и закономерное поведение материи, основанное не только на одной тенденции переходить от упорядоченности к неупорядоченности, но и частично на существовании упорядоченности, которая поддерживается все время... — ... средство, при помощи которого организм поддерживает себя постоянно на достаточно высоком уровне упорядоченности (равно на достаточно низком уровне энтропии), в действительности состоит в непрерывном извлечении упорядоченности из окружающей среды».
У высших животных нам хорошо известен тот вид упорядоченности, которым они питаются, а именно: крайне упорядоченное состояние материи в более или менее сложных органических соединениях служит им пищей. После использования животные возвращают эти вещества в очень деградированной форме, однако, не вполне деградированной, так как их еще могут усваивать растения.
Для растений мощным источником «отрицательной энтропии» — негэнтропии - является солнечный свет.
Свойство живых систем извлекать упорядоченность из окружающей среды дало основание некоторым ученым сделать вывод, что для этих систем второе начало термодинамики не выполняется. Однако второе начало имеет еще и другую, более общую формулировку, справедливую для открытых, в том числе живых, систем. Она гласит, что эффективность самопроизвольного превращения энергии всегда меньше 100%. В соответствии со вторым началом термодинамики поддержание жизни на Земле без притока солнечной энергии невозможно.
Обратимся снова к Э. Шредингеру: «Все, что происходит в природе, означает увеличение энтропии в той части Вселенной, где это имеет место. Так и живой организм непрерывно увеличивает свою энтропию, или производит положительную энтропию и, таким образом, приближается к опасному состоянию — максимальной энтропии, представляющему собой смерть. Он может избежать этого состояния, т.е. оставаться живым, только постоянно извлекая из окружающей среды отрицательную энтропию».
Перенос энергии в экосистемах и ее потери
Как известно, в перенос энергии пищи от ее источника — растений — через ряд организмов, происходящий путем поедания одних организмов другими, проходит через пищевую цепь. При каждом очередном переносе большая часть (80-90%) потенциальной энергии теряется, переходя в тепло. Переход к каждому следующему звену уменьшает доступную энергию примерно в 10 раз. Экологическая энергетическая пирамида всегда сужается кверху, поскольку энергия на каждом последующем уровне теряется (рис. 1).
Эффективность природных систем много ниже КПД электромоторов и других двигателей. В живых системах много «горючего» уходит на «ремонт», что не учитывается при расчете КПД двигателей. Любое повышение эффективности биологической системы оборачивается увеличением затрат на их поддержание в устойчивом состоянии. Экологическую систему можно сравнить с машиной, из которой нельзя «выжать» больше, чем она способна дать. Всегда наступает предел, после которого выигрыш от роста эффективности сводится на нет ростом расходов и риском разрушения системы. Прямое удаление человеком или животными более 30-50% годового прироста растительности может уменьшить способность экосистемы сопротивляться стрессу.
Один из пределов биосферы — валовая продукция фотосинтеза, и под него человеку придется подгонять свои нужды, пока не удастся доказать, что усвоение энергии путем фотосинтеза можно сильно повысить, не подвергая при этом опасности нарушить равновесие других, более важных ресурсов жизненного круговорота. Сейчас же поглощается лишь около половины всей лучистой энергии (в основном, в видимой части спектра) и, самое большее, — около 5% — ее в самых благоприятных условиях превращается в продукт фотосинтеза.

Рис. 1. Пирамида энергий. Е — энергия, выделяемая с метаболитами; D = естественные смерти; W — фекалии; R — дыхание
В искусственных экосистемах для получения большего урожая человек вынужден расходовать добавочную энергию. Она необходима для индустриализованного сельского хозяйства, так как этого требуют культуры, специально созданные для него. «Индустриализованное (использующее энергию горючих ископаемых) сельское хозяйство (как, например, практикуемое в Японии) может дать в 4 раза более высокий урожай с гектара, чем сельское хозяйство, в котором всю работу выполняют люди и домашние животные (как в Индии), но оно требует в 10 раз больших затрат разного рода ресурсов и энергии».
Замкнутость производственных циклов по энергетически энтропийному параметру теоретически невозможно, поскольку течение энергетических процессов (в соответствии со вторым началом термодинамики) сопровождается деградацией энергии и повышением энтропии природной среды. Действие второго начала термодинамики выражается в том, что превращения энергии идут в одном направлении в отличие от цикличного движения веществ.
В настоящее время мы являемся свидетелями того, что повышение уровня организации и разнообразия культурной системы уменьшает ее энтропию, но увеличивает энтропию окружающей природной среды, вызывая ее деградацию. В какой степени можно элиминировать эти следствия второго начала термодинамики? Существуют два пути.
Первый путь заключается в уменьшении потерь используемой человеком энергии при ее различных превращениях. Этот путь эффективен в той мере, в которой не приводит к понижению стабильности системы, через которую идет поток энергии (как известно, в экологических системах увеличение числа трофических уровней способствует повышению их устойчивости, но в то же время способствует росту потерь энергии, проходящей через систему).
Второй путь заключается в переходе от повышения упорядоченности культурной системы к повышению упорядоченности всей биосферы. Общество в этом случае повышает организованность природной среды за счет понижения организованности части той природы, которая находится за пределами биосферы Земли.
Превращение веществ и энергии в биосфере как открытой системе
Принципиальное значение для понимания динамики биосферных процессов и конструктивного решения конкретных экологических проблем имеют теория и методы открытых систем, являющиеся одним из важнейших достижений XX столетия.
Согласно классической теории термодинамике, физические и другие системы неживой природы эволюционируют в направлении усиления их беспорядка, разрушения и дезорганизации. При этом энергетическая мера неорганизованности, выраженная энтропией, имеет тенденцию к непрерывному увеличению. Возникает вопрос: каким же образом из неживой природы, системы которой имеют тенденцию к дезорганизации, могла появиться живая природа, системы которой в своей эволюции стремятся к совершенствованию и усложнению своей организации? К тому же, в обществе в целом прогресс очевиден. Следовательно, исходное понятие классической физики — понятие закрытой или изолированной системы не отражает реальности и находится в явном противоречии с результатами исследований в биологии и общественных науках (например, мрачные прогнозы «тепловой смерти» Вселенной). И вполне закономерно, что в 1960-е годы появляется новая (нелинейная) термодинамика, основывающаяся на концепции необратимых процессов. Место закрытой, изолированной системы в ней занимает принципиально иное основополагающее понятие открытой системы, которая способна обмениваться с окружающей средой веществом, энергией и информацией. Средство, с помощью которого организм поддерживает себя на достаточно высоком уровне упорядоченности (равно на достаточно низком уровне энтропии), в действительности состоит в непрерывном извлечении упорядоченности из окружающей среды.
Открытая система , таким образом, заимствует извне либо новое вещество, либо свежую энергию и одновременно выводит во внешнюю среду использованное вещество и отработанную энергию, т.е. она не может оставаться замкнутой. В процессе эволюции система постоянно обменивается энергией с окружающей средой и производит энтропию. При этом характеризующая степень беспорядка в системе энтропия, в отличие от закрытых систем, не аккумулируется, а транспортируется в окружающую среду. Логичен вывод, что открытая система не может быть равновесной , поскольку требует непрерывного поступления из внешней среды энергии или богатого ею вещества. По Э. Шредингеру, вследствие такого взаимодействия система черпает из окружающей среды порядок и тем самым привносит в нее беспорядок.
Взаимодействие между экосистемами
Если между двумя системами существует связь, возможен переход энтропии из одной системы в другую, вектор которого определяется значениями термодинамических потенциалов. Здесь-то и проявляется качественное различие между изолированными и открытыми системами. В изолированной системе ситуация остается неравновесной. Процессы идут, пока энтропия не достигнет максимума.
В открытых системах отток энтропии наружу может уравновесить ее рост в самой системе. Такого рода условия способствуют возникновению и поддержанию стационарного состояния (типа динамического равновесия), называемого текущим равновесием. В стационарном состоянии энтропия открытой системы остается постоянной, хотя и не является максимальной. Постоянство поддерживается за счет того, что система непрерывно извлекает из окружающей среды свободную энергию.
Динамика энтропии в открытой системе описывается уравнением И.Р. Пригожина (бельгийский физик, лауреат Нобелевской премии 1977 г.):
ds/dt = ds 1 /dt + ds e /dt,
где ds 1 /dt - характеристика энтропии необратимых процессов внутри самой системы; ds e /dt - характеристика обмена энтропией между биологической системой и окружающей средой.
Саморегулирование флуктуирующих экосистем
Суммарное уменьшение энтропии в результате обмена с внешней средой при определенных условиях может превысить ее внутреннее производство. Появляется неустойчивость предшествующего неупорядоченного состояния. Возникают и возрастают до макроскопического уровня крупномасштабные флуктуации. При этом возможна саморегуляция , т.е. возникновение определенных структур из хаотических образований. Такие структуры могут последовательно переходить во все более упорядоченное состояние (дис- сипативные структуры). Энтропия в них убывает.
Диссипативные структуры образуются вследствие развития собственных внутренних неустойчивостей в системе (в результате самоорганизации), что отличает их от организации упорядоченных структур, формирующихся под воздействием внешних причин.
Упорядоченные (диссипативные) структуры, спонтанно возникающие из беспорядка и хаоса в результате процесса самоорганизации, реализуются и в экологических системах. Примером может служить пространственно упорядоченное расположение бактерий в питательных средах, наблюдающееся при определенных условиях, а также временные структуры в системе «хищник-жертва», отличающиеся устойчивым режимом колебаний с определенной периодичностью численности популяций животных.
Процессы самоорганизации основываются на обмене энергией и массой с окружающей средой. Это и позволяет поддерживать искусственно создаваемое состояние текущего равновесия, когда потери на диссипацию компенсируются извне. С поступлением новой энергии или вещества в системе возрастает неравновесность. В конечном итоге прежние взаимосвязи между элементами системы, определяющие ее структуру, разрушаются. Между элементами системы устанавливаются новые связи, приводящие к кооперативным процессам, т.е. к коллективному поведению ее элементов. Такова общая схема процессов самоорганизации в открытых системах, названная наукой синергетикой .
Концепция самоорганизации, по-новому освещая взаимосвязь неживой и живой природы, позволяет лучше понять, что весь окружающий нас мир и Вселенная представляют собой совокупность самоорганизующихся процессов, которые лежат в основе любого эволюционного развития.
Целесообразно обратить внимание на следующее обстоятельство. Исходя из случайного характера флуктуации следует, что появление нового в мире всегда обусловлено действием случайных факторов.
Возникновение самоорганизации опирается на принцип положительной обратной связи, в соответствии с которым изменения, возникающие в системе, не устраняются, а накапливаются. В итоге именно это и приводит к возникновению нового порядка и новой структуры.
Точка бифуркации — импульс развития биосферы по новому пути
Открытые системы физической Вселенной (к которым относится и наша биосфера) непрерывно флуктуируют и на определенном этапе могут достигнуть точки бифуркации . Суть бифуркации наиболее наглядно иллюстрирует сказочный витязь, стоящий на распутье. В каком-то месте пути встречается развилка, где необходимо принимать решение. При достижении точки бифуркации принципиально нельзя предугадать, в каком направлении будет дальше развиваться система: перейдет ли она в хаотическое состояние или приобретет новый, более высокий уровень организации.
Для точка бифуркации — импульс к ее развитию по новому, неведомому пути. Какое место займет в нем человеческое общество — предугадать сложно, биосфера же, наиболее вероятно, продолжит свое развитие.
Энтропия представляет меру вероятности пребывания системы в данном состоянии. Оказалось, что энтропия является одним из фундаментальных свойств любых систем с вероятностным поведением, обеспечивающим новые уровни понимания в кодировании информации, в системном анализе, лингвистике, биологии, обработке изображений и т. п. Влияние внешней информации на систему может быть оценено через изменение энтропии состояния системы.
При достижении системой стационарного состояния суммарное изменение энтропии можно считать приблизительно равным нулю, что соответствует взаимной компенсации всех процессов, связанных с поступлением, удалением и превращением вещества, энергии и информации.
Нобелевский лауреат И. Пригожин сформулировал основное свойство стационарного состояния открытых систем при фиксированных внешних параметрах скорость воспроизведения энтропии, обусловленная протеканием необратимых процессов, постоянна во времени и минимальная по величине dЭ/dt → min.
Таким образом, согласно теореме И. Пригожина, стационарное состояние характеризуется минимальным рассеянием энтропии. Для живых систем это положение можно сформулировать так: поддержание гомеостазиса требует минимального потребления энергии, т.е. здоровый организм стремится работать в самом экономном энергетическом режиме. Что касается заболеваний организма, то оно связано с дополнительными энергетическими затратами для компенсации приобретенных или врожденных биологических дефектов и с ростом энтропии.
В динамической системе может быть несколько стационарных состояний, отличающихся уровнем воспроизведения энтропии dЭk/dt. С этой точки зрения состояние организма может быть описано в виде набора энергетических уровней, некоторые из которых устойчивы (уровни 1 и 4), другие нестабильные (уровни 2, 3, 5). При наличии постоянно действующего внешнего или внутреннего возмущения может происходить скачкообразный переход из одного состояния в другое. Любое восполнение характеризуется увеличенным потреблением энергии: температура тела повышается, увеличивается скорость обменных процессов. Отклонение от стационарного состояния с минимальными энергозатратами вызывает развитие внутренних процессов, стремящихся вернуть систему обратно, к уровню 1. При длительных действиях факторов система может перейти к уровню 3, в так называемую точку бифуркации, из которой возможно несколько исходов: возвращение на стабильный уровень 1, переход в другое устойчивое равновесное состояние 4, характеризующееся новым энергоинформационным уровнем, или скачок на более высокий, нестабильный уровень 5.
Для организма это соответствует нескольким адаптационным уровням относительно здоровья или хронического заболевания с разными уровнями функционирования системы. Острое заболевание соответствует нестационарному состоянию с повышенным воспроизведением энтропии, т.е. неэкономному типу функционирования организма. Согласно математической теории катастроф, при острых заболеваниях необходимо скачком перевести организм из "плохого" устойчивого состояния в "хорошее". При этом используют большие дозы лекарственных препаратов. В фазе затухающего обострения болезней возрастает роль малых воздействий, например акупунктуры и гомеопатических средств, оказывающих положительное воздействие.
Мультистабильность сложных нелинейных систем, какой является организм человека, вероятностная природа его постоянного развития и самоорганизация приводят к необходимости поиска "системообразующих факторов", к которым относится и энтропия.
Как показано в, энтропия является информативной характеристикой состояния организма и может использоваться для оценки его состояния и определения направления терапии больных. В предложен метод вычисления энтропии газоразрядной визуализации (ГРВ) биологических объектов и соответствующее программное обеспечение.
«Не может человек найти суть дела, что делается под солнцем,
-сколько б ни трудился искать человек – не найдет;
и если даже скажет мудрец, что сумеет, - не найдет».
Соломон Мудрый, царь иудейский, Х век до н.э.
Таков сей мир, а отчего он так,
Того не ведает ни умный, ни дурак.
Д. И. Фонвизин (1745 – 1792).
Системой можно назвать совокупность взаимодействующих частей. Опытным фактом является то обстоятельство, что некоторые свойства частей диктуются самой системой, что интегративные, системные свойства этой совокупности не являются свойствами самих частей. Для человека с индуктивным мышлением эта идея является крамолой и ее хочется предать анафеме.
Клетка в живом организме человека.
Клетка человека является частью организма. Внутренний геометрический объем клетки ограничен от внешней среды мембраной, оболочкой. Через эту границу происходит взаимодействие среды и клетки. Будем рассматривать клетку человека с ее оболочкой как термодинамическую систему, даже если великие термодинамики современности сочтут клетку собственного организма вульгарным и недостойным для термодинамики объектом рассмотрения.
По отношению к клетке человека внешняя среда – это межклеточная жидкость, водный раствор. Ее состав определяется обменом химическими веществами с кровеносными сосудами (капиллярами) и обменом с множеством клеток. Из межклеточной жидкости в клетку через оболочку поступают «полезные» вещества и кислород. Из клетки через ту же оболочку выходят в межклеточную жидкость продукты жизнедеятельности, это необходимые для организма вещества, побочные продукты, шлаки, не прореагировавшие компоненты. Следовательно, клетка человека, как термодинамическая система, взаимодействует с внешней средой химически . Потенциал этого взаимодействия обозначим традиционно буквой μ, а координату состояния этого рода взаимодействия обозначим m. Тогда количество этого взаимодействия внешнего мира и клетки организма равно
где j – номер маршрута последовательных и/или параллельных химических превращений, m j – масса вновь образовавшегося j-го вещества. Индекс (е) вверху означает, что следует брать величину j-го потенциала превращения для внешней среды, т.е. для межклеточной жидкости.
Одновременно, через оболочку клетки организма осуществляется термическое взаимодействие с потенциалом Т (абсолютная температура) и координатой термического рода s (энтропия). Количество взаимодействия – T (e) ds.
Деформационным взаимодействием (потенциал – давление, координата состояния – удельный объем системы) для жидкостей пренебрегаем.
Тогда первый закон термодинамики для термо-химической системы записывается в стандартной форме:
du = μ j (e) dm j + T (e) ds ,
где u – внутренняя энергия системы.
Если потенциалы в клетке организма μ j (i) и Т (i) близки к потенциалам снаружи, то наступает равновесие. Равновесность означает, что количество исходных реагентов и количество продуктов реакций в обратимых химических превращениях становятся неизменными (все химические реакции – обратимы).
Системное свойство организма заключается в том, что функциональное назначение каждой клетки человека - производство веществ, необходимых организму (белки, жиры, ферменты, энергоносители и т.п.). Клетка должна выдавать эти вещества в межклеточную жидкость и далее в кровеносную систему. Следовательно, состояние клетки человека должно быть неравновесным, а процессы обмена – необратимыми. Это значит, что если
Δμ j = μ j (e) – μ j (i) , то Δμ j /μ j (i) ≥ 10 0 .
Для рассматриваемой ситуации (необратимость) первый закон термодинамики принимает вид:
du = T (e) ds + (Δμ j + μ j (i))dm j = T (e) ds + μ j (i) dm j + Δμ j dm j .
Последний член в этом уравнении обусловлен необратимостью процесса химического взаимодействия. И, согласно второму закону термодинамики, эта необратимость обязательно приводит к росту энтропии:
Δμ j dm j = T (i) ds (m) дисс, где ds (m) дисс > 0. (дисс = диссипация).
Все происходит так, как будто необратимость при взаимодействии любого рода «включает» в термодинамической системе источник теплоты с активностью T (i) ds (m) дисс, клетка организма нагревается (не обязательно в смысле роста температуры, как на кухне, а в более широком смысле - подвода теплоты). Рост энтропии в клетке человека безусловно искажает течение химических реакций (об этом чуть дальше). Происходит генерация ненужных организму веществ, мусора, шлаков, происходит разбавление раствора. Организму приходится отводить энтропию из клетки, а то она такого ему наделает!
Один из путей отвода энтропии указывает термодинамика: необходимо уменьшить термический потенциал Т (е) , сделать его меньше Т (i) . А чтобы реализовать отвод теплоты, разность температур ΔТ = Т (i) – Т (e) должна снова быть конечной величиной, следовательно, процесс теплообмена станет тоже необратимым, появится еще один источник теплоты с активностью T (i) ds (T) дисс. Окончательно, первый закон термодинамики для термо-химической системы с необратимыми процессами обмена примет вид:
du = T (i) ds + μ j (i) dm j + T (i) ds (m) дисс + Т (i) ds (T) дисс.
Первые два члена в du справа ответственны за обратимые процессы взаимодействия, последние два – за необратимые, причем последний обусловлен предпоследним. Следовательно, часть внутренней энергии системы необратимо превращается в теплоту, т.е. клетка человека генерирует энтропию .
На этом остановимся в применении термодинамического метода анализа клетки в живом организме. Остановка определяется смыслом эпиграфов к этой статье: для этого метода исследования необходима еще количественная информация, которой мы не располагаем. Но и то, что получили, дорогого стоит! Осталось делать комментарий и получать следствия.
Чем опасна энтропия в клетке организма?
Попробуем понять, почему рост энтропии ds (m) дисс > 0 и ds (T) дисс > 0 опасен для организма. А может быть этот рост благоприятен?
Организм «требует» от клетки ее функционирования, выполнения полезных и необходимых ему потребительских услуг в виде производства каких-то веществ. Причем, требует реализации этих услуг «быстро» в каком-то смысле. Скорость превращений обусловлена конечностью разностей потенциалов, применением катализаторов и специальных транспортных молекул. Но в любой ситуации необходимо расположить молекулы реагентов плотно и рядом (в геометрическом смысле). Далее, молекулы реагентов за счет своей энергии Е должны «возбудить» электронные оболочки каких-то атомов, затем может произойти акт соединения, синтеза с образованием новых веществ.
Молекулы в клетке человека имеют, как правило, сложную пространственную трехмерную структуру. И потому у таких молекул много степеней свободы движения элементов. Это может быть вращательное движение фрагментов молекулы, это может быть колебательное движение тех же фрагментов и отдельных атомов. Наверное, вращение крупных фрагментов молекулы в жидкой фазе затруднено, очень уж тесно. Вращаются, по-видимому, только мелкие фрагменты. А вот колебаниям мелких фрагментов и отдельных атомов молекулы высокая плотность жидкой фазы не очень мешает. Во всяком случае, число степеней свободы движения у такой молекулы огромно, следовательно, общее число W вариантов распределения энергии Е по этим степеням свободы еще больше. Если следовать Больцману и принять
то рост энтропии в клетке организма приводит к отводу энергии от вариантов, способных возбудить электронные оболочки с последующим образованием «нужных» веществ. Да еще при таком росте энтропии начинают синтезироваться побочные продукты.
Организму придется навести порядок в клетке человека, отвести энтропию из объема клетки, чтобы сосредоточить энергию молекул в «полезных» степенях свободы. Бедный организм, даже на клеточном уровне у него нет халявы: хочешь что-то ценное получить, убирай энтропию из клетки.
Методы интенсификации отвода энтропии.
Из теории теплопередачи следует, что количество теплоты
dQ = kF(T (i) – T (e)) dτ = (T (i) ds (m) дисс + T (i) ds (T) дисс)ρV,
где k – коэффициент теплопередачи, F – поверхность теплообмена (оболочки клетки организма), τ – время, ρ – плотность системы. Поделим обе части этого уравнения на объем клетки V. Тогда слева появится множитель F/V ∼ d -1 , где d – характерный размер клетки организма. Следовательно, чем мельче клетка, тем интенсивнее идет процесс отвода энтропии при одной и той же разности термических потенциалов. Более того, с уменьшением размера d можно уменьшить эту разность при том же dQ и, следовательно, меру термической необратимости ds (T) дисс.
Иными словами, генерация энтропии происходит в объеме клетки V ∼ d 3 , а отвод энтропии из клетки человека осуществляется через поверхность F ∼ d 2 (см. рис. 1).
Рис. 1. Иллюстрация к определению критического размера клетки организма.
Но клетка увеличивает свою массу и, следовательно, объем. И пока d d 0 поверхность отводит меньше энтропии, чем ее генерируется, да еще в темпе внешней среды. При d > d 0 , наступит «разогрев» клетки, она начнет вредить организму. Что же делать? С одной стороны, клетке человека надо увеличивать свою массу, а, с другой, нельзя увеличивать свой размер. Единственный путь «спасения» клетки и организма – деление клетки. Из «большой» клетки размером d 0 (считая пока для простоты клетку человека сферической) образуется два «ребенка» размером d р:
πd 0 3 /6 = 2πd 3 р /6 > d р = 2 -1/3 d 0 = 0,794d 0 .
Размер «детей» будет на 20% меньше размера «матушки». На рис. 2 представлена динамика размера клетки человека в организме.

Рис. 2. Динамика размера клетки организма. d 00 – размер клетки у новорожденного.
Замечание . Увеличение интенсивности отвода энтропии из клетки человека возможно не только уменьшением температуры T (e) межклеточной жидкости и, следовательно, крови в капиллярах, но и увеличением температуры T (i) внутри клетки организма. Но такой способ изменит весь химизм в клетке, она перестанет выполнять свои функции в организме, да еще начнет производить всякий «мусор». Вспомните, как вам плохо из-за высокой температуры при каком-то заболевании. Температуру в клетке человека лучше не трогать, для работоспособности с позиции организма клетке придется регулярно делиться, и это же обстоятельство уменьшает прирост ds (T) дисс > 0.
Еще одно замечание . Если рассмотреть удельную поверхность тел различной геометрической формы, то не сложно увидеть, что минимальная удельная поверхность у шара. Поэтому на Севере и в Сибири жители строят дома в виде полусфер, да еще стараются делать дома большими по размеру (d > d 0) на 2-3 семьи. Это позволяет существенно экономить свои силы на заготовку дров к зиме. Зато в жарких странах дома строят в виде вытянутых тел с большим числом пристроек. Для интенсификации отвода энтропии из клетки человека последняя должна иметь форму, далекую от шара.
Энтропия управляет всем.
Теперь попробуем представить себе, что было бы, если бы делились еще и нервные клетки человека (нейроны со своими отростками-дендритами и синапсами на их конце). Нейрофизиолог сразу пришел бы в ужас от такой перспективы: это означало бы просто разрушение всей системы иннервации организма и работы мозга. Только-только человек усвоил какое-то знание, приобрел какой-то навык, прием, и вдруг все исчезло, начинай снова или пропадай.
Простым аналогом деления нервных клеток являются путчи, смуты, бунты и революции, т.е. смена команды правящей элиты в какой-то стране. А народы потом долго корчатся, приспосабливаясь к новым правителям. Нет, чисто функционально нервным клеткам человека нельзя позволить делиться!
Как же это реализуется, ведь энтропия в клетках организма неумолимо растет? Прежде всего, обратим внимание на разветвленность нервной клетки человека, на большое развитие ее теплообменной поверхности (поверхность тонкой длинной нити много больше поверхности шара того же объема).
Далее, оказывается, организм тщательно бдит за температурой артериальной крови, поступающей в головной мозг. Проявляется это, в частности, в том, что у теплокровных животных создана автономная система (малый круг) кровообращения. Единственный температурный датчик находится в сонной артерии, с помощью его организм управляет температурой артериальной крови, приходящей в мозг. Забота о регулировании этой температуры дошла до того, что теплокровные наземные животные имеют дополнительную возможность охлаждения крови, поступающей в мозг. Оказывается, сонная артерия разветвляется так, что часть крови по байпасу проходит через ушные раковины-теплообменники. Специальный датчик управляет расходом этой крови. Если температура увеличилась сверх номинальной, то этот расход увеличивается, кровь остывает в ушах на ветерке, далее смешивается с основным потоком и направляется в мозг.
Вспомните бедного африканского слона: в жару приходится все время махать ушами. Вспомните, какие большие уши у млекопитающих в жарких странах, и какие маленькие в холодных. В русской бане, в парной следует закрывать именно уши, чтобы подольше с удовольствием попариться. На лыжной прогулке зимой опять-таки надо закрывать уши, чтобы не охлаждать мозг. У студента-двоечника, мечтающего о позорной тройке, на экзамене или зачете уши всегда красные, а у отличника – уши нормального цвета. По цвету ушей можно сразу определять оценку!
Ну, а когда человеческая головка совсем перестала соображать, т.е. накопила многовато энтропии в нервных клетках головного мозга, то придется пойти погулять, сменить вид деятельности, например, порубить дрова. Наконец, просто поспать, снять нагрузку на нейроны головного мозга, уменьшить производство энтропии и за 8 часов ночного сна вывести ее из головного мозга с помощью венозной крови. Получается, что накопление энтропии в нервных клетках человека определяет весь режим его жизни: с утра едем на работу, потом едем домой с работы, немного отдыха и далее сон.
Вот бы придумать такой механизм отвода энтропии из нервных клеток, чтобы можно было все 24 часа в сутки работать! Сколько было бы радости для творческих людей и для эксплуататоров! ВВП в стране вырос бы сразу более чем на 30%! Не нужен транспорт для перевозки людей, не нужны жилища, а только рабочие места. Организация жизни стала бы простейшей: ребенок непрерывно учится в школе, потом в институте или профтехучилище, далее человек помещается на рабочем месте и в конце отвозится в крематорий. Фантасты, хватай идею!
Наверное, понятно, что производство разных целевых продуктов для организма приводит к разной интенсивности генерации энтропии в разных клетках человека. Все определяется «сложностью», т.е. пространственной архитектурой молекул целевого вещества и разнообразием и числом радикалов и атомов в ее составе. Чем больше эта «сложность», тем больше уменьшается энтропия при синтезе из простых радикалов, но и тем больше прирост диссипативной энтропии.
Производство мужских половых гормонов у теплокровных наземных животных отличается от производства других необходимых организму веществ. Суть дела в том, что в этом гормоне должно быть огромное количество информации, которое организм - папа хочет передать женской яйцеклетке. Он озабочен передачей своих свойств и черт своему ребенку, так как они позволили папе выживать в окружающем его макромире.
Специалисты в теории информации утверждают, что информация без ее материальных носителей не существует. И таким носителем информации о свойствах и чертах папы является молекула гормона, точнее, ее архитектура, набор и расположение фрагментов, радикалов и атомов элементов из таблицы Д.И. Менделеева. И чем больше количество информации, чем она подробнее и детальнее, тем сложнее молекула гормона. Шаг вправо, шаг влево – образуется мутация, отклонение от мечтаний папы. Следовательно, синтез такой молекулы означает существенное уменьшение энтропии в системе, и одновременно производство в клетке человека еще большего количества диссипативной энтропии.
Простой аналогией является строительство здания. Строительство царского Зимнего дворца в Петербурге со всеми его архитектурными излишествами и роскошью означает сильное уменьшение энтропии по сравнению со строительством деревенских изб той же полезной площади, но зато количество мусора (энтропии) после завершения – несоизмеримо.
Производство мужских половых гормонов у теплокровных наземных животных так интенсивно генерирует диссипативную энтропию, что межклеточная жидкость с кровеносными сосудами не может столько ее отвести из клеток. Бедному самцу пришлось выделить эти органы наружу для обдува холодным атмосферным воздухом. Если молодой парень сидит на скамейке в метро или в автобусе, широко раздвинув колени к вящему возмущению соседок-старушек, то не обвиняйте его в хамстве, это выходит энтропия. А мальчишки в возрасте до 15 лет, старички и женщины всех возрастов сидят, скромно и культурно сдвинув колени.
И в женской яйцеклетке после ее образования происходят химические превращения, поддерживающие ее в «боеспособном» состоянии. Но энтропия неумолимо увеличивается со временем, отвода теплоты по существу нет, приходится организму выбрасывать яйцеклетку, а потом делать новую, создавая массу неприятностей нашим милым дамам. Если этого не делать, то или зачатия не будет, или будут рождаться всякие ужастики. У других млекопитающих этих проблем с энтропией в яйцеклетке нет, они готовы к деторождению в течение малого промежутка времени, да еще строго дискретно: слоны – раз в 5–6 лет, человекообразные обезьяны – раз в 3 года, коровы – раз в год, кошки – 3–4 раза в год. Зато человек – практически непрерывно. И за что его так отяготила природа? А, может быть, осчастливила? Тайна!
Рассмотрена одна из наиболее важных термодинамических функций - энтропия. Описаны свойства этой функции и значение, которое она имеет в биосистемах.
ЭНТРОПИЯ БИОСИСТЕМ
ВВЕДЕНИЕ
Среди термодинамических функций, характеризующих энергетическое состояние биологического объекта, исключительно важное место принадлежит энтропии. Понятие энтропии было введено в 1865 году Рудольфом Клаузиусом, и с тех пор эта функция привлекает внимание физиков и физикохимиков. Широко используется энтропия и биологами , что вполне понятно. Ведь живой организм - это прежде всего энергетическая система, где действуют те же законы термодинамики, что и в неживой природе. Следует, однако, учесть, что живые организмы характеризуются некоторыми особенностями, которые отсутствуют у физических объектов. Это, как известно, размножение, развитие и т.д. Поэтому энергетический обмен таких систем обладает качественным своеобразием и требует специального анализа.
Почему для этого анализа мы взяли именно энтропию? Это связано с тем, что данная функция дает наиболее полную и в то же время обобщенную характеристику системы. Она может указывать, какие процессы возможны в данных условиях и до какого предела они могут идти.
ЧТО ТАКОЕ ЭНТРОПИЯ
Энтропия - очень "скользкая" термодинамическая функция. Многие о ней слышали, но далеко не все представляют, что это такое. Конечно, можно сказать (как это сделает физик), что изменение энтропии системы при равновесном процессе равно отношению теплоты, сообщенной системе, к абсолютной температуре: dS = dQ / T, что с повышением температуры (неадиабатическим путем) энтропия системы возрастает, что она представляет собой функцию состояния, то есть ее изменение не зависит от формы пути и, следовательно, в замкнутом контуре ее изменение равно нулю и т.д. Но все это мало удовлетворит биолога, поэтому начнем несколько издалека.
В биосистемах протекают самые разные энергетические процессы: дыхание, фотосинтез, мышечное сокращение, транспорт веществ и т.д. Однако при всем качественном разнообразии эти процессы можно попытаться свести к двум типам: обратимые и необратимые. Обратимый процесс - это такой, при котором система в каждый данный момент времени находится в состоянии, бесконечно близком к термодинамическому равновесию, и достаточно лишь незначительно изменить условия, чтобы процесс был обращен. При этом под термодинамическим равновесием понимают такое состояние системы, когда градиенты различных видов энергии (химической, электрической) выровнены и способность системы совершать работу равна нулю. Обращение обратимого процесса не вызывает остаточных изменений в окружающей среде.
В отличие от этого при необратимом процессе система изменяется по направлению к конечному состоянию (при самопроизвольном протекании процесса - к состоянию равновесия) с определенной скоростью. При этом часть свободной энергии системы (то есть той энергии системы, за счет которой может совершаться работа при постоянной температуре) теряется в виде тепла. Например, если совершается механическая работа, то часть затраченной на нее свободной энергии теряется при трении. Для того чтобы обратить данный процесс, необходимо компенсировать эти потери. Следовательно, обращение необратимого процесса связано с остаточными изменениями в окружающей среде. Потери энергии при необратимых процессах, которые происходят обычно в виде тепла, и характеризует энтропия. Таким образом, энтропия отражает ту часть энергии системы, которая рассеялась, деградировала в тепловой форме и не может уже быть использована для совершения работы при постоянной температуре. Из сказанного становится ясным, что при обратимых процессах изменение энтропии равно нулю (DS = 0), а при необратимых оно положительно (DS > 0). Таким образом, чем меньше в системе градиенты энергии и чем больше в ней рассеянной в виде тепла деградированной энергии, тем больше ее энтропия.
Особенностью биосистем является то, что в них практически нет обратимых процессов. Все процессы, которые в них протекают, носят необратимый характер, то есть сопровождаются увеличением энтропии. Следовательно, в биосистемах не вся затрачиваемая при данном процессе свободная энергия переходит в полезную работу. Часть ее рассеивается в виде тепла. Отношение количества совершенной работы к количеству затраченной на нее свободной энергии называется коэффициентом полезного действия биологического процесса. Так, мышечное сокращение совершается с КПД ~ 30%, гликолиз ~ 36% и т.д. Как видим, потери свободной энергии при этих процессах весьма велики. Встречаются, однако, и такие процессы, которые близки к обратимым, то есть КПД которых высок. Например, свечение некоторых тропических насекомых имеет КПД 98-99%, разряд электрических рыб - 98%. Причина такого высокоэффективного использования свободной энергии пока не совсем ясна. Таким образом, мы приходим к выводу, что, чем больше увеличение энтропии при данном процессе, тем более он необратим.
БИОСИСТЕМЫ И ВТОРОЙ ПРИНЦИП ТЕРМОДИНАМИКИ
Значение энтропии особенно четко проявляется при рассмотрении второго принципа термодинамики . Этот принцип, как известно, представляет собой фундаментальный закон природы и в общей форме устанавливает, что в изолированной системе энергия самопроизвольно может переходить только от более высокого уровня к более низкому, а не наоборот. Как постулировал Р. Клаузиус, "теплота не может переходить сама собой от более холодного тела к более теплому".
Пользуясь понятием энтропии, можно придать второму принципу более конкретный вид: в изолированной системе могут самопроизвольно протекать только такие процессы, при которых энтропия или остается постоянной (обратимые), или увеличивается (необратимые). Самопроизвольного уменьшения энтропии в такой системе не происходит.
Применим ли второй принцип термодинамики к биосистемам? Ответ на этот вопрос не так прост. В биосистемах протекают процессы, при которых энергия в соответствии с этим принципом переходит с более высокого на более низкий уровень. Это, например, процесс дыхания. В ходе его богатые энергией соединения (углеводы) распадаются до простых низкоэнергетических веществ - воды и углекислоты, а выделившаяся свободная энергия используется для протекания других процессов (например, синтеза АТФ). Однако хорошо известно, что в живых системах осуществляются и такие процессы, в ходе которых энергия переходит с более низкого на более высокий уровень. Так, например, происходит при фотосинтезе. Здесь, как известно, из простых бедных энергией соединений углекислоты и воды при участии квантов света синтезируются вещества (например, углеводы), содержащие значительный запас свободной энергии. Можно назвать и другие процессы в биосистемах, протекание которых, казалось бы, не подчиняется второму принципу термодинамики. Это позволило некоторым ученым говорить о том, что данный принцип не действует в биосистемах.
Но так ли это? Ответ на этот вопрос связан с выбором системы для рассмотрения. Необходимо рассматривать организм не изолированно от внешней среды, а во взаимодействии с ней. Как известно, в изолированной системе самопроизвольно протекающие процессы приводят ее к состоянию так называемого термодинамического равновесия. При этом состоянии градиенты энергии выровнены, работоспособность системы равна нулю, а ее энтропия максимальна. Однако хорошо известно, что биологические системы, пока они функционируют, никогда не достигают такого состояния. Они всегда обладают работоспособностью, а их энтропия немаксимальна. Это связано с тем, что биосистемы являются не изолированными, а открытыми системами.